Aberrant Hypomethylation of Solute Carrier Family 6 Member 12 Promoter Induces Metastasis of Ovarian Cancer
- Affiliations
-
- 1Department of Biochemistry, School of Medicine, Ewha Womans University, Seoul, Korea. ahnj@ewha.ac.kr
- 2Department of Biomedical Sciences, Seoul National University College of Medicine, Seoul, Korea.
- 3College of Pharmacy and Research Institute of Life and Pharmaceutical Sciences, Sunchon National University, Suncheon, Korea.
- 4Department of Obstetrics and Gynecology, School of Medicine, Ewha Womans University, Seoul, Korea. goodmorning@ewha.ac.kr
- KMID: 2374185
- DOI: http://doi.org/10.3349/ymj.2017.58.1.27
Abstract
- PURPOSE
Ovarian cancer (OC) is the most fatal of gynecological malignancies with a high rate of recurrence. We aimed to evaluate the expression of solute carrier family 6, member 12 (SLC6A12) and methylation of its promoter CpG sites in a xenograft mouse model of metastatic OC, and to investigate the regulatory mechanisms that promote aggressive properties during OC progression.
MATERIALS AND METHODS
Expression of SLC6A12 mRNA was determined by reverse-transcription quantitative polymerase chain reaction (RT-qPCR), and DNA methylation status of its promoter CpGs was detected by quantitative methylation-specific PCR. The metastatic potential of SLC6A12 was evaluated by in vitro migration/invasion transwell assays. Gene expression and DNA methylation of SLC6A12 and clinical outcomes were further investigated from publicly available databases from curatedOvarianData and The Cancer Genome Atlas.
RESULTS
SLC6A12 expression was 8.1-14.0-fold upregulated and its DNA methylation of promoter CpG sites was 41-62% decreased in tumor metastases. After treatment with DNA methyltransferase inhibitor and/or histone deacetylase inhibitor, the expression of SLC6A12 was profoundly enhanced (~8.0-fold), strongly supporting DNA methylation-dependent epigenetic regulation of SLC6A12. Overexpression of SLC6A12 led to increased migration and invasion of ovarian carcinoma cells in vitro, approximately 2.0-fold and 3.3-fold, respectively. The meta-analysis showed that high expression of SLC6A12 was significantly associated with poor overall survival [hazard ratio (HR)=1.07, p value=0.016] and that low DNA methylation levels of SLC6A12 at specific promoter CpG site negatively affected patient survival.
CONCLUSION
Our findings provide novel evidence for the biological and clinical significance of SLC6A12 as a metastasis-promoting gene.
Keyword
MeSH Terms
-
Animals
Carrier Proteins/genetics/*metabolism
Cell Line, Tumor
Cell Migration Assays
*CpG Islands
*DNA Methylation
Disease Progression
Epigenesis, Genetic
Female
Gene Expression Regulation, Neoplastic
Humans
Mice
Neoplasm Invasiveness
Neoplasm Transplantation
Ovarian Neoplasms/genetics/*metabolism/mortality/pathology
Polymerase Chain Reaction
Prognosis
*Promoter Regions, Genetic
RNA, Messenger/*metabolism
Up-Regulation
Carrier Proteins
RNA, Messenger
Figure
-
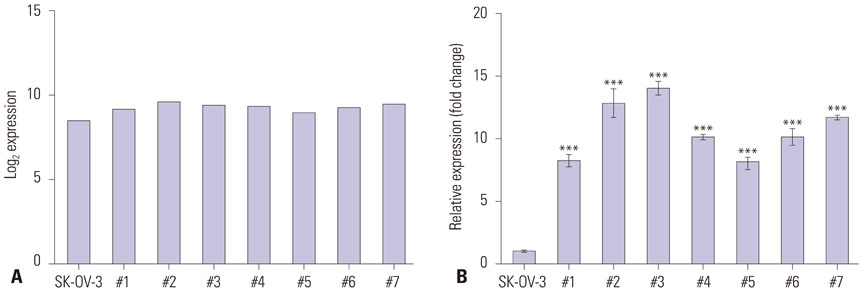
Fig. 1 Expression of SLC6A12 mRNA was upregulated in tumor metastases harvested from ovarian cancer xenograft mice. The expression of SLC6A12 mRNA was measured by expression microarray (A) and RT-qPCR (B). The error bars indicate means±standard deviation (SD) of triplicate experiments. Statistical analyses were performed using one-way analysis of variance (ANOVA) and Turkey’s multiple comparison post-test for comparing the significance between mouse metastases with SK-OV-3 cells (***p<0.001). The tumor metastases from each ovarian cancer xenograft mouse were labeled #1–#7 (n=7). SLC6A12, solute carrier family 6, member 12; RT-qPCR, reverse-transcription quantitative polymerase chain reaction.
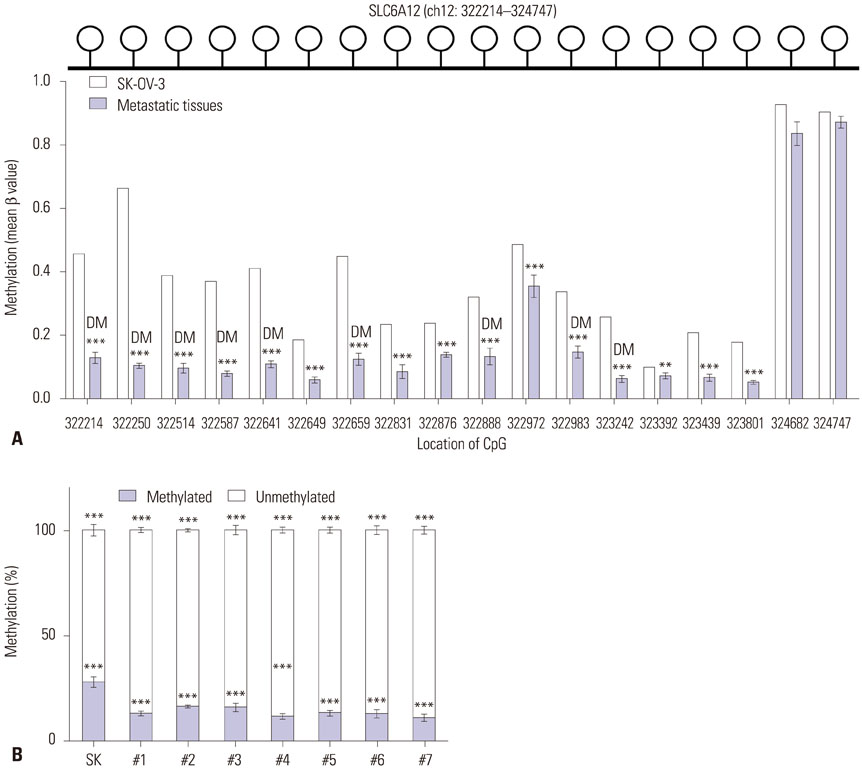
Fig. 2 Alteration of DNA methylation of the CpG sites of SLC6A12 in tumor metastases from ovarian cancer xenograft mice. The DNA methylation status of the SLC6A12 promoter region was analyzed using the Illumina HumanMethylation450 BeadChip, which contains 18 specific CpG sites at positions 322214–324747 (the human GRCh37/hg19 assembly) within chromosome 12 (upper panel of A). Individual bars represent the mean β-value of methylation at the corresponding CpG site within the SLC6A12 promoter (lower panel of A). Data are shown as the mean±SD. Statistical analyses were performed using Bayesian t-statistics (**p<0.01, ***p<0.001) (A). Differences in the DNA methylation status of the 323242 CpG site in SK-OV-3 cells and tumor metastases was analyzed with qMSP. Statistical analyses were performed using one-way ANOVA and Turkey’s multiple comparison post-tests for comparing significance with SK-OV-3 cells (***p<0.001) (B). DM, differentially methylated CpG site; SLC6A12, solute carrier family 6, member 12; qMSP, quantitative methylation-specific polymerase chain reaction.

Fig. 3 Modulation of SLC6A12 mRNA expression following demethylation in SK-OV-3 cells. SK-OV-3 cells were treated for 72 h with various concentration of 5-aza-2’-deoxycytidine (5-aza-dC). After treatment, expression of SLC6A12 mRNA was measured by RT-qPCR (A), and the methylation status at the 323242 CpG site was analyzed with qMSP (B). Alternatively, SK-OV-3 cells were treated with 10 µM of 5-aza-dC for 72 h and then with/without 0.5 µM of trichostatin (TSA) for 24 h. After treatment, expression of SLC6A12 mRNA was measured by RT-qPCR (C) and the methylation status of the 323242 CpG site was analyzed with qMSP (D). Data are shown as the mean±SD (n=3). Statistical analyses were performed using one-way ANOVA with Turkey’s multiple comparison post-test for comparing significance with untreated control or 5-aza-dC/TSA treated samples (*p<0.05, **p<0.01, ***p<0.001). SLC6A12, solute carrier family 6, member 12; RT-qPCR, reverse-transcription quantitative polymerase chain reaction; qMSP, quantitative methylation-specific polymerase chain reaction.
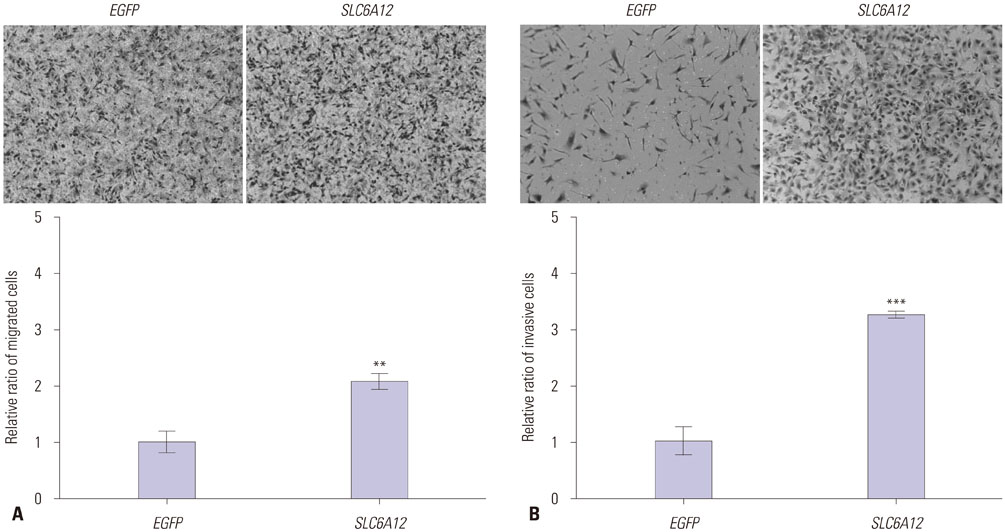
Fig. 4 Overexpression of SLC6A12 enhanced migration and invasiveness of SK-OV-3 cells. Migration of serum-starved cells towards 15% serum-containing medium was determined by transwell assay. Representative images of migrated cells transfected with EGFP or SLC6A12 are shown. Quantitative analysis of migrated cells was carried out by measuring the absorbance of stained cell extracts at 595 nm (200×, magnification) (A). Migration of serum-starved cells towards 10% serum-containing medium was determined using a Matrigel-coated invasion chamber. Representative images of invading cells transfected with EGFP or SLC6A12 are shown. Quantitative analysis of invading cells was carried out by measuring the absorbance of stained cell extracts at 595 nm (200×, magnification) (B). Data are shown as the mean±SD for triplicate measurements. Statistical analysis was performed using a t-test (**p<0.01, ***p<0.001). SLC6A12, solute carrier family 6, member 12.
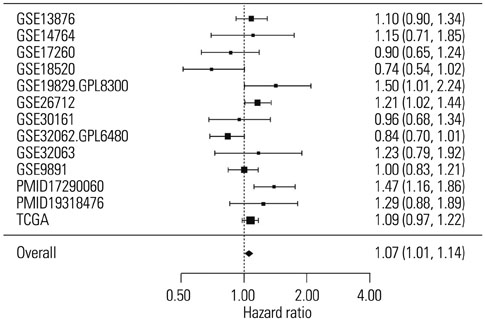
Fig. 5 Increased expression of SLC6A12 was associated with a high risk of death in ovarian cancer patients. Forest plot of the significant association between increased risk of death and high expression of SLC6A12 in 1979 serous-type ovarian cancer patients from 13 independent studies found in curatedOvarianData. SLC6A12, solute carrier family 6, member 12.
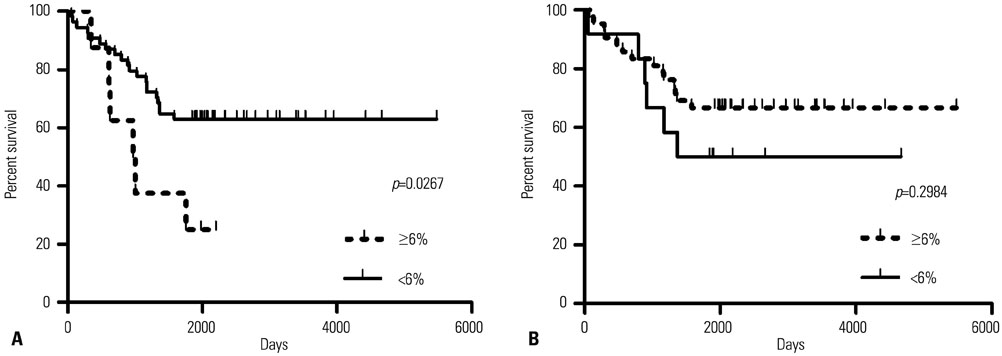
Fig. 6 The survival of ovarian cancer patients was affected by the methylation status of SLC6A12. Kaplan-Meier analysis of overall survival in patients with serous ovarian cancer. The 5-year survival of patients associated with methylation status of SLC6A12 of <6% methylation versus ≥6% methylation of the 323242 CpG site (A) and 322250 CpG site (B) was compared and the statistical significance between two groups was determined by univariate Cox regression analysis. SLC6A12, solute carrier family 6, member 12.
Reference
-
1. American Cancer Society. Cancer facts & figures 2015. Atlanta (GA): American Cancer Society;2015.2. Lengyel E. Ovarian cancer development and metastasis. Am J Pathol. 2010; 177:1053–1064.
Article3. Asadollahi R, Hyde CA, Zhong XY. Epigenetics of ovarian cancer: from the lab to the clinic. Gynecol Oncol. 2010; 118:81–87.
Article4. Balch C, Fang F, Matei DE, Huang TH, Nephew KP. Minireview: epigenetic changes in ovarian cancer. Endocrinology. 2009; 150:4003–4011.
Article5. Zhou Y, Holmseth S, Hua R, Lehre AC, Olofsson AM, Poblete-Naredo I, et al. The betaine-GABA transporter (BGT1, SLC6A12) is predominantly expressed in the liver and at lower levels in the kidneys and at the brain surface. Am J Physiol Renal Physiol. 2012; 302:F316–F328.6. Kempson SA, Zhou Y, Danbolt NC. The betaine/GABA transporter and betaine: roles in brain, kidney, and liver. Front Physiol. 2014; 5:159.
Article7. Smyth GK. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat Appl Genet Mol Biol. 2004; 3:Article3.
Article8. Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Series B Stat Methodol. 1995; 289–300.
Article9. Kim NH, Sung HY, Choi EN, Lyu D, Choi HJ, Ju W, et al. Aberrant DNA methylation in the IFITM1 promoter enhances the metastatic phenotype in an intraperitoneal xenograft model of human ovarian cancer. Oncol Rep. 2014; 31:2139–2146.
Article10. Ganzfried BF, Riester M, Haibe-Kains B, Risch T, Tyekucheva S, Jazic I, et al. curatedOvarianData: clinically annotated data for the ovarian cancer transcriptome. Database (Oxford). 2013; 2013:bat013.
Article11. Reimers M, Carey VJ. Bioconductor: an open source framework for bioinformatics and computational biology. Methods Enzymol. 2006; 411:119–134.12. Dugué AE, Pulido M, Chabaud S, Belin L, Gal J. How to deal with interval-censored data practically while assessing the progression-free survival: a step-by-step guide using SAS and R software. Clin Cancer Res. 2016; 09. 12. [Epub]. DOI: 10.1158/1078-0432.CCR-16-1017.
Article13. Zhang D, Li X, Yao Z, Wei C, Ning N, Li J. GABAergic signaling facilitates breast cancer metastasis by promoting ERK1/2-dependent phosphorylation. Cancer Lett. 2014; 348:100–108.
Article14. Wu W, Yang Q, Fung KM, Humphreys MR, Brame LS, Cao A, et al. Linking γ-aminobutyric acid A receptor to epidermal growth factor receptor pathways activation in human prostate cancer. Mol Cell Endocrinol. 2014; 383:69–79.
Article15. Sung HY, Choi EN, Lyu D, Park AK, Ju W, Ahn JH. Aberrant hypomethylation-mediated AGR2 overexpression induces an aggressive phenotype in ovarian cancer cells. Oncol Rep. 2014; 32:815–820.
Article16. Sung HY, Ju W, Ahn JH. DNA hypomethylation-mediated overexpression of carbonic anhydrase 9 induces an aggressive phenotype in ovarian cancer cells. Yonsei Med J. 2014; 55:1656–1663.
Article17. Sung HY, Park AK, Ju W, Ahn JH. Overexpression of mucin 13 due to promoter methylation promotes aggressive behavior in ovarian cancer cells. Yonsei Med J. 2014; 55:1206–1213.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- DNA Hypomethylation-Mediated Overexpression of Carbonic Anhydrase 9 Induces an Aggressive Phenotype in Ovarian Cancer Cells
- The Role of the SLC Transporters Protein in the Neurodegenerative Disorders
- Clinical Study of a Newly Diagnosed Case of Gitelman Syndrome in a Patient Monitored for Liddle Syndrome
- Overexpression of Mucin 13 due to Promoter Methylation Promotes Aggressive Behavior in Ovarian Cancer Cells
- Epigenetic Changes (Aberrant DNA Methylation) in Colorectal Neoplasia
