Calcium/Calmodulin-Dependent Protein Kinase is Involved in the Release of High Mobility Group Box 1 Via the Interferon-beta Signaling Pathway
- Affiliations
-
- 1Department of Pathology, Hallym University College of Medicine, Chuncheon 200-702, Korea. kwonik@hallym.ac.kr
Abstract
- Previously, we have reported that high mobility group box 1 (HMGB1), a proinflammatory mediator in sepsis, is released via the IFN-beta-mediated JAK/STAT pathway. However, detailed mechanisms are still unclear. In this study, we dissected upstream signaling pathways of HMGB1 release using various molecular biology methods. Here, we found that calcium/calmodulin-dependent protein kinase (CaM kinase, CaMK) is involved in HMGB1 release by regulating IFN-beta production. CaMK inhibitor, STO609, treatment inhibits LPS-induced IFN-beta production, which is correlated with the phosphorylation of interferon regulatory factor 3 (IRF3). Additionally, we show that CaMK-I plays a major role in IFN-beta production although other CaMK members also seem to contribute to this event. Furthermore, the CaMK inhibitor treatment reduced IFN-beta production in a murine endotoxemia. Our results suggest CaMKs contribute to HMGB1 release by enhancing IFN-beta production in sepsis.
Keyword
MeSH Terms
-
Benzimidazoles
Cytokines
Endotoxemia
HMGB1 Protein
Inflammation
Interferon Regulatory Factor-3
Interferon-beta
Molecular Biology
Naphthalimides
Phosphorylation
Phosphotransferases
Protein Kinases
Sepsis
Signal Transduction
Benzimidazoles
Cytokines
HMGB1 Protein
Interferon Regulatory Factor-3
Interferon-beta
Naphthalimides
Phosphotransferases
Protein Kinases
Figure
-
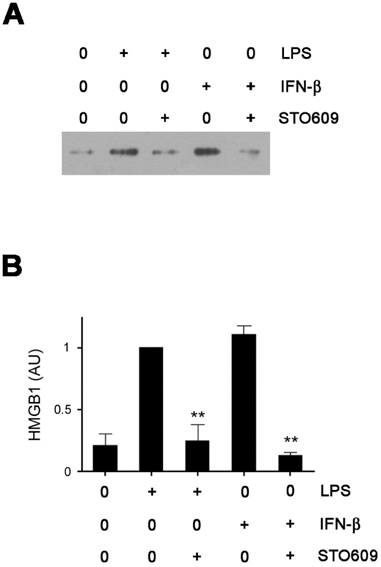
Figure 1 CaMKs are indispensable to HMGB1 secretion. (A) RAW cells were pre-incubated with STO609 for 30 minutes and then treated with LPS (1 mg/ml) or IFN-β (500 U/ml) overnight. The levels of HMGB1 in supernatants were measured by western blotting. (B) The relative densitometric values of HMGB1 bands are shown as arbitrary units (AU). Densitometric value from the LPS-treated cells was taken as 1 and the mean values±SEM fold changes relative to the control (the LPS-treated cells) are shown (n=3). **p<0.01 compared with LPS (or IFN-β)-treated control samples.
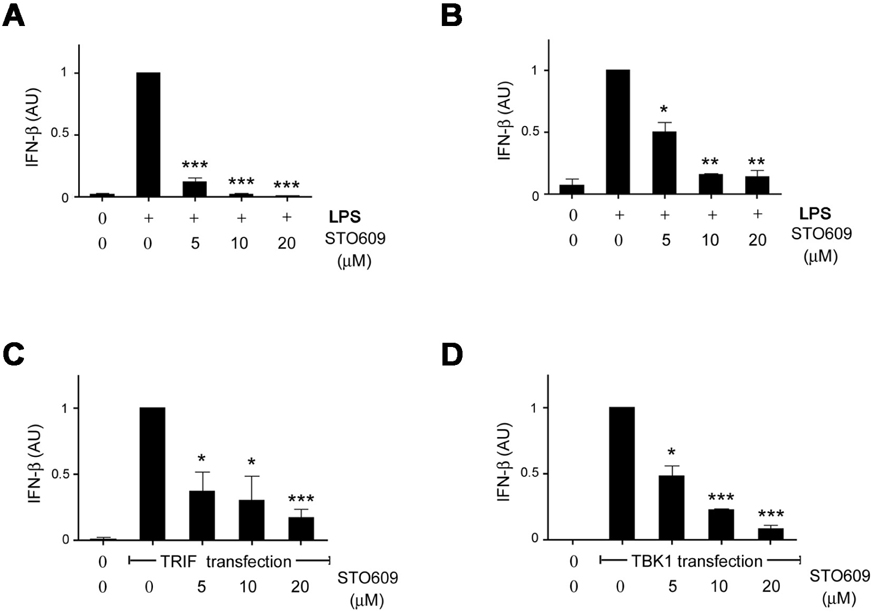
Figure 2 Inhibition of CaMKs blocks IFN-β production. (A and B) CaMK inhibitor STO609 inhibits IFN-β production in LPS-treated RAW cells (A) and macrophages (B) in a dose-dependent manner (n=3). RAW cells or peritoneal macrophages were stimulated with LPS (1 µg/ml) in the presence of vehicle (DMSO) or various doses of STO609 for 2 hours. The mRNA were extracted from each cells and subjected to real time PCR. The value of the LPS-treated cells was arbitrarily assigned a value of 1. (C and D) 293 cells were transfected with TRIF (C) or TBK1 (D) plasmids. After 24 hours of culture, IFN-β production was measured by real time PCR and the normalized values of IFN-β were analyzed the same as in A (n=3). Data were expressed as mean±SEM. AU, arbitrary unit. *p<0.05; **p<0.01; ***p<0.001 compared with each control.

Figure 3 Inhibition of CaMK interferes with IRF3 phosphorylation induced by LPS. (A) STO609 treatment reduced IRF3 phosphorylation. RAW cells were treated with LPS in the presence of various doses of STO609 for 1 hour, lysed and then subjected to phospho-IRF3 western blotting (middle). The same blot was subjected again to HMGB1 western blotting as a loading control. A graph shows the relative densitometric values of each sample in arbitrary unit (AU). Densitometirc value from the LPS-treated cells was taken as 1. (B) TBK1 phosphorylation was not affected by STO609. Raw cells were treated with LPS plus STO609 and the cell lysates were immunoprecipitated with anti-phospho-TBK1 antibody and then detected with anti-total TBK1 antibody (middle). Lower panel, equal amounts of each lysate were subjected to HMGB1 western blotting as a loading control. **p<0.01; ***p<0.001 compared with LPS alone.
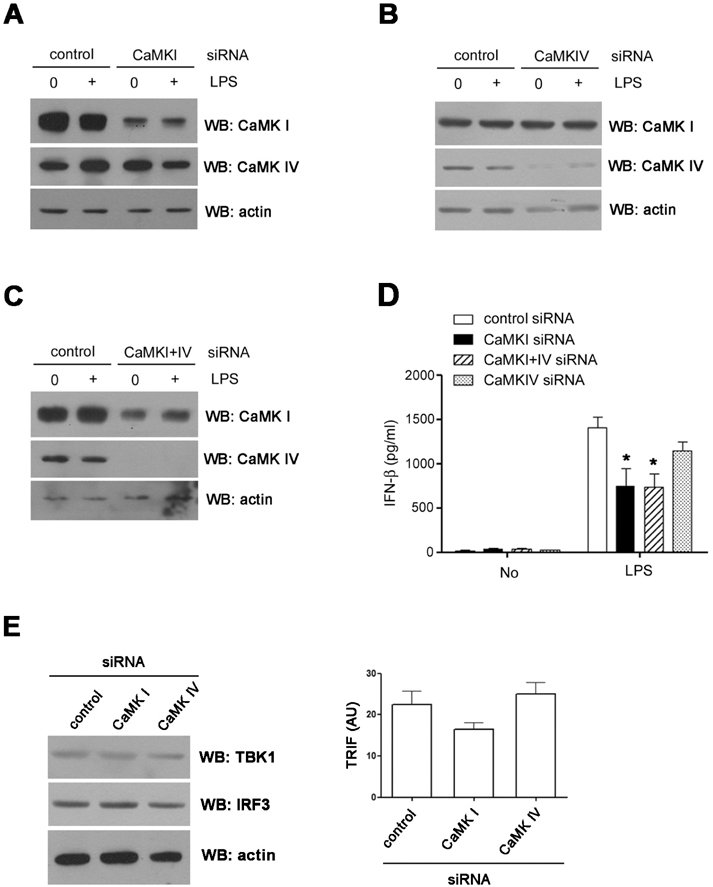
Figure 4 CaMK1 plays a role in IFN-β production. (A~C) RAW cells were transfected with CaMKI (A), IV (B) or I and IV (C) siRNAs. After 48 hours, the expressions of each CaMK isoform were detected by western blotting. (D) The levels of IFN-β in the supernatant were measured by ELISA. (E) The levels of TBK1, IRF3 proteins (left) and TRIF transcripts (right) in siRNA treated cells were analyzed using western blotting (left) or real time PCR. *p<0.05 compared with LPS treated cells transfected with control siRNA.

Figure 5 The release of IFN-β depends on CaMK signaling pathway in vivo. (A and B) Serum levels of IFN-β (A) and TNF (B) in B6 mice treated with LPS (50 mg/kg) plus various doses of STO609 or vehicle (DMSO) were measured by ELISA (n=4). B6 mice were treated various doses of STO609 twice (18 and 0.5 hours) before LPS infusion. Mice were sacrificed 2 hours after LPS administration and sera were collected from each mouse for ELISA. Data are presented as mean±SEM. *p<0.05 compared with LPS plus vehicle-treated mice.
Reference
-
1. Sims GP, Rowe DC, Rietdijk ST, Herbst R, Coyle AJ. HMGB1 and RAGE in inflammation and cancer. Annu Rev Immunol. 2010. 28:367–388.
Article2. Lotze MT, Tracey KJ. High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal. Nat Rev Immunol. 2005. 5:331–342.
Article3. Degryse B, Bonaldi T, Scaffidi P, Müller S, Resnati M, Sanvito F, Arrigoni G, Bianchi ME. The high mobility group (HMG) boxes of the nuclear protein HMG1 induce chemotaxis and cytoskeleton reorganization in rat smooth muscle cells. J Cell Biol. 2001. 152:1197–1206.
Article4. Rouhiainen A, Kuja-Panula J, Wilkman E, Pakkanen J, Stenfors J, Tuominen RK, Lepäntalo M, Carpén O, Parkkinen J, Rauvala H. Regulation of monocyte migration by amphoterin (HMGB1). Blood. 2004. 104:1174–1182.
Article5. Sappington PL, Yang R, Yang H, Tracey KJ, Delude RL, Fink MP. HMGB1 B box increases the permeability of Caco-2 enterocytic monolayers and impairs intestinal barrier function in mice. Gastroenterology. 2002. 123:790–802.
Article6. Andersson U, Wang H, Palmblad K, Aveberger AC, Bloom O, Erlandsson-Harris H, Janson A, Kokkola R, Zhang M, Yang H, Tracey KJ. High mobility group 1 protein (HMG-1) stimulates proinflammatory cytokine synthesis in human monocytes. J Exp Med. 2000. 192:565–570.
Article7. Youn JH, Oh YJ, Kim ES, Choi JE, Shin JS. High mobility group box 1 protein binding to lipopolysaccharide facilitates transfer of lipopolysaccharide to CD14 and enhances lipopolysaccharide-mediated TNF-alpha production in human monocytes. J Immunol. 2008. 180:5067–5074.
Article8. Sha Y, Zmijewski J, Xu Z, Abraham E. HMGB1 develops enhanced proinflammatory activity by binding to cytokines. J Immunol. 2008. 180:2531–2537.
Article9. Bonaldi T, Talamo F, Scaffidi P, Ferrera D, Porto A, Bachi A, Rubartelli A, Agresti A, Bianchi ME. Monocytic cells hyperacetylate chromatin protein HMGB1 to redirect it towards secretion. EMBO J. 2003. 22:5551–5560.
Article10. Kim JH, Kim SJ, Lee IS, Lee MS, Uematsu S, Akira S, Oh KI. Bacterial endotoxin induces the release of high mobility group box 1 via the IFN-beta signaling pathway. J Immunol. 2009. 182:2458–2466.
Article11. Zhang X, Wheeler D, Tang Y, Guo L, Shapiro RA, Ribar TJ, Means AR, Billiar TR, Angus DC, Rosengart MR. Calcium/calmodulin-dependent protein kinase (CaMK) IV mediates nucleocytoplasmic shuttling and release of HMGB1 during lipopolysaccharide stimulation of macrophages. J Immunol. 2008. 181:5015–5023.
Article12. Wang L, Tassiulas I, Park-Min KH, Reid AC, Gil-Henn H, Schlessinger J, Baron R, Zhang JJ, Ivashkiv LB. 'Tuning' of type I interferon-induced Jak-STAT1 signaling by calcium-dependent kinases in macrophages. Nat Immunol. 2008. 9:186–193.
Article13. Tokumitsu H, Inuzuka H, Ishikawa Y, Ikeda M, Saji I, Kobayashi R. STO-609, a specific inhibitor of the Ca(2+)/calmodulin-dependent protein kinase kinase. J Biol Chem. 2002. 277:15813–15818.
Article14. Karaghiosoff M, Steinborn R, Kovarik P, Kriegshäuser G, Baccarini M, Donabauer B, Reichart U, Kolbe T, Bogdan C, Leanderson T, Levy D, Decker T, Müller M. Central role for type I interferons and Tyk2 in lipopolysaccharide-induced endotoxin shock. Nat Immunol. 2003. 4:471–477.
Article15. Weighardt H, Kaiser-Moore S, Schlautkötter S, Rossmann-Bloeck T, Schleicher U, Bogdan C, Holzmann B. Type I IFN modulates host defense and late hyperinflammation in septic peritonitis. J Immunol. 2006. 177:5623–5630.
Article16. Liu X, Yao M, Li N, Wang C, Zheng Y, Cao X. CaMKII promotes TLR-triggered proinflammatory cytokine and type I interferon production by directly binding and activating TAK1 and IRF3 in macrophages. Blood. 2008. 112:4961–4970.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Role of Calmodulin in the Generation of Reactive Oxygen Species and Apoptosis Induced by Tamoxifen in HepG2 Human Hepatoma Cells
- PKR as a Regulator of Inflammasome Activation
- Ras Mitogen-activated Protein Kinase Signaling and Kinase Suppressor of Ras as Therapeutic Targets for Hepatocellular Carcinoma
- A Memory Molecule, Ca2+/Calmodulin-Dependent Protein Kinase II and Redox Stress; Key Factors for Arrhythmias in a Diseased Heart
- Regulation of Protein Kinase in KCl-induced Contraction of Cat Gastric Smooth Muscle
