Hypoxia Activates Toll-like Receptor 4 Signaling in Primary Mouse Hepatocytes Through the Receptor Clustering within Lipid Rafts
- Affiliations
-
- 1Department of Surgery, Eulji General Hospital, Eulji University School of Medicine, Seoul, Korea. kdh2109@eulji.ac.kr
- 2Department of Surgery, University of Pittsburgh, Pittsburgh, PA 15213, US.
- KMID: 1963583
- DOI: http://doi.org/10.4174/jkss.2011.80.3.194
Abstract
- PURPOSE
Transient hypoxia is an initial event that accentuates ischemia/reperfusion (I/R) injury in the liver. Hepatic ischemia/reperfusion (I/R) injury is largely related to innate immunity via Toll-like receptor 4 (TLR4) signaling. However, the mechanism by which hypoxia could lead to activate TLR4 signaling remains unclear. Therefore, the aim of this experimental study investigates how TLR4 signalling is activated by hypoxia.
METHODS
Hepatocytes were isolated from male wild-type (C57BL/6) mice (8~12 weeks old) by an in situ collagenase (Type IV, Sigma-Aldrich) perfusion technique. In this study, using primary mouse hepatocytes in culture to 1% oxygen, detection of TLR4 translocation to the lipid rafts on the cell membrane by immunofluorescence staining and immunoblotting was saught.
RESULTS
Hypoxia caused TLR4/MD2 and beta2-Integrin (CD11b/CD18) translocation to lipid rafts associated with CD14 in hepatocytes. The cholesterol sequestering agent, Nystatin and Filipin prevented hypoxia-induced TLR4/MD2 translocation to lipid rafts. Consistent with a role for oxidative stress in this effect, in vitro H2O2 treatment of hepatocytes similarly caused TLR4/MD2 translocation to lipid rafts. In addition, translocation of hypoxia-induced TLR4 complex was inhibited by N-acetylcysteine (NAC) demonstrating that the activation of TLR4 signaling is dependent on ROS. Further, the cholesterol sequestering agent, nystatin, prevented hypoxia-induced high mobility group box 1 (HMGB1) release in hepatocytes.
CONCLUSION
These results suggest that ROS dependent TLR4 signaling is achieved following receptor translocation to the lipid raft in hepatocytes. We hypothesized that this mechanism is required for the release of HMGB1, an early mediator of injury and inflammation in hepatic I/R injury.
Keyword
MeSH Terms
-
Acetylcysteine
Animals
Anoxia*
Cell Membrane
Cholesterol
Cluster Analysis*
Collagenases
Filipin
Fluorescent Antibody Technique
Hepatocytes*
HMGB1 Protein
Humans
Immunity, Innate
Immunoblotting
Inflammation
Liver
Male
Mice*
Nystatin
Oxidative Stress
Oxygen
Perfusion
Sequestering Agents
Toll-Like Receptor 4*
Acetylcysteine
Cholesterol
Collagenases
Filipin
HMGB1 Protein
Nystatin
Oxygen
Sequestering Agents
Toll-Like Receptor 4
Figure
-
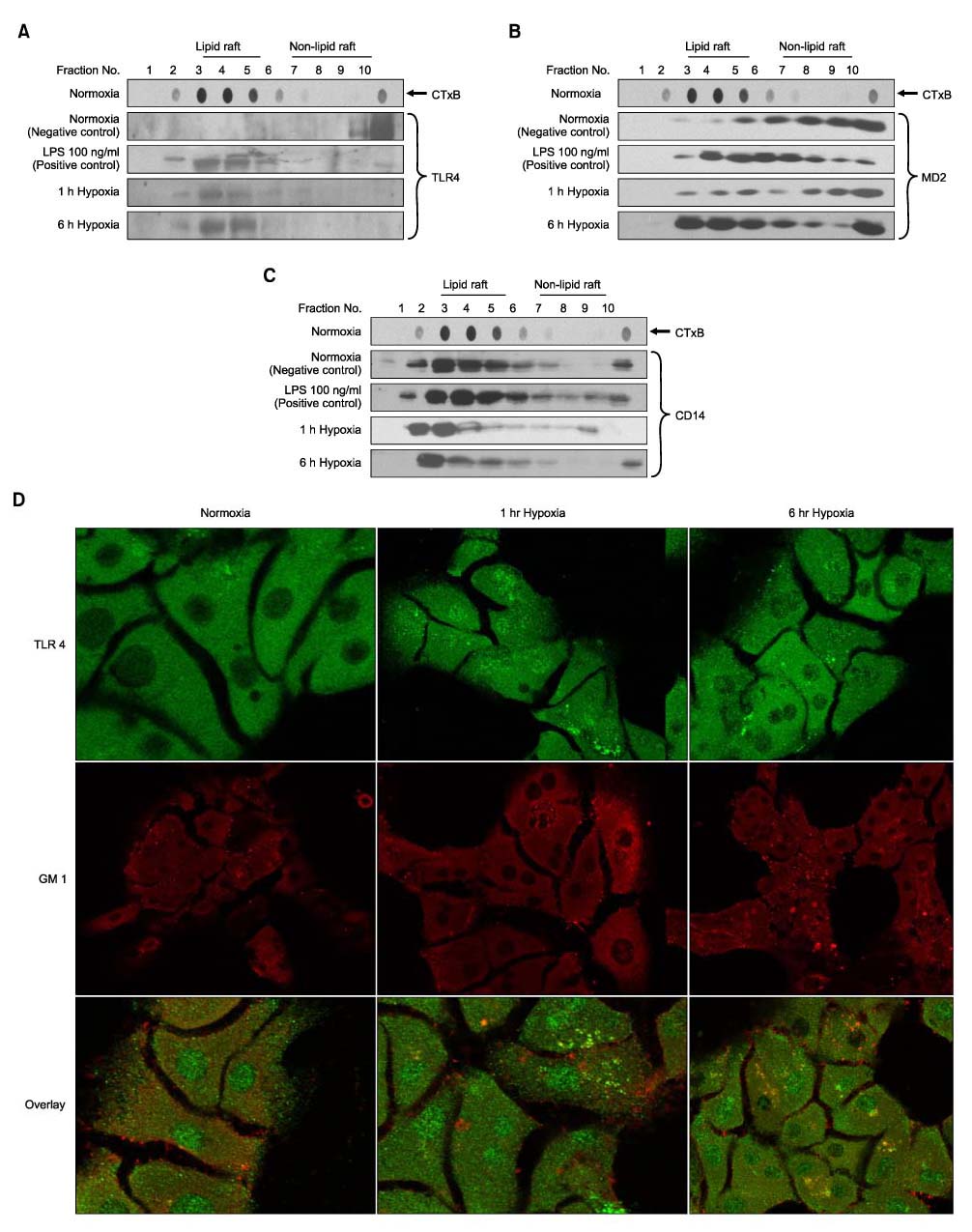
Fig. 1 Hypoxia causes TLR4/MD2 complex translocation to lipid rafts associated with CD14. Distribution and localization of TLR4, MD2 and CD14 on the membrane in mouse hepatocyes. Cells were cultured under normoxic (21% oxygen) and hypoxic (1% oxygen) conditions and in the presence or absence of LPS stimulation. Each fraction was obtained using discontinuous sucrose gradient ultracentrifugation. Initially, each fraction was analyzed by dot blotting using CTxB conjugated to horseradish peroxidase. Fractions 3 to 5 were designated as lipid raft as indicated by CTxB (GM1, top row). And then equal aliquots of each fraction were run on a polyacrylamide gel, blotted and probed with antibody to TLR4, MD2 and CD14 and peroxidase-coupled secondary antibody. TLR4 and MD2 translocated to lipid rafts after LPS stimulation, 1 or 6-hour hypoxia. CD-14 was found in lipid rafts regardless of normoxia or hypoxia. The blot pattern for TLR4 (A), MD2 (B), and CD14 (C) represents three individual experiments. Blot shown is representative of three different experiments with similar results. (D) TLR4 and GM1 colocalization after hypoxia. Hepatocytes were plated on coverslips and then fixed, permeabilized at 4℃. The cells were stained with anti-TLR4 primary as well as the FITC conjugated corresponding secondary antibodies or rhodamine-CTxB. Representative images are shown of TLR4 (green), rhodamine-CTxB staining (red), or the merged image (yellow).
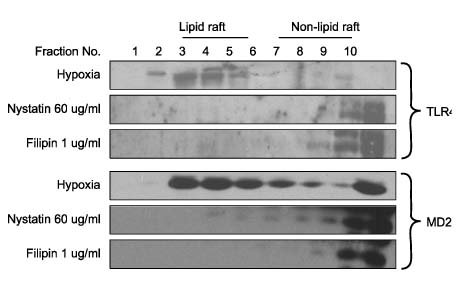
Fig. 2 Disruption of lipid raft layer blocks hypoxia induced TLR4/MD2 complex translocation. nystatin (60µg/ml) or filipin (1µg/ml) inhibits hypoxia-induced TLR4/MD2 translocation to lipid raft. Hepatocytes were pre-incubated with nystatin (60µg/ml) or filipin (1µg/ml) for 30 min or 1 hour before exposure of hypoxia. After centrifugation, 1 ml fractions were collected from the top. Equal aliquots of each fraction were run on polyacrylamide gels, blotted and probed with TLR4 or MD2 antibody.
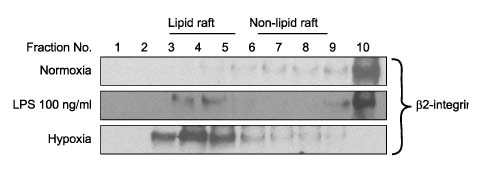
Fig. 3 Hypoxia causes β2-Integrin translocation to lipid rafts. Distribution and localization of β2-Integrin (CD11b/CD18) on the membrane in mouse hepatocytes. Cells were cultured under normoxic (21% oxygen) and hypoxic (1% oxygen) conditions and in the presence or absence of LPS stimulation. Each fraction was obtained using discontinuous sucrose gradient ultracentrifugation. Equal aliquots of each fraction were run on a polyacrylamide gel, blotted and probed with antibody to β2-Integrin (CD11b/CD18) and peroxidase-coupled secondary antibody. β2-Integrin (CD11b/CD18) translocated to lipid rafts after LPS stimulation or 6-hour hypoxia.
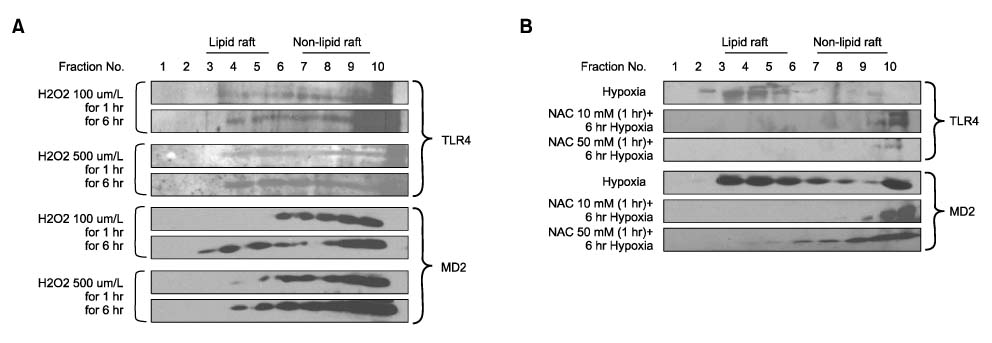
Fig. 4 The TLR4/MD2 complex translocation is ROS dependent signaling. (A) Hepatocytes were treated with H2O2 in concentration from 50~500µM for 1 or 6 hours and subjected to discontinuous sucrose density gradient centrifugation as described in materials and methods. Equal aliquots of each fraction were run on a polyacrylamide gel, blotted and probed with antibody to TLR4 or MD2 and peroxidase-coupled secondary antibody. The H2O2 was shown to induce a dose and time-dependent increased TLR4/MD2 translocation to lipid rafts. (B) Hepatocytes were treated with N-acethylcystein (NAC) in concentration from 10 mM or 50 mM for 1hour before exposure to hypoxia (1% oxygen). NAC blocked both TLR4 and MD2 translocation to lipid rafts.
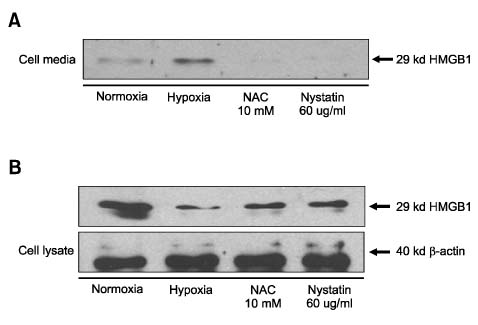
Fig. 5 Disruption of lipid raft layer blocks hypoxia induced HMGB1 release. Hepatocytes were pre-incubated with nystatin (60µg/ml) or N-acethylcystein (10 mM) for 30 min or 1 hour. The cells were then cultured under hypoxic (1% oxygen) conditions for 24 hours. The media from cells were concentrated with filter device and then supernatants were collected. Western blot analysis for HMGB1 in supernatants (A) and in cell lysates (B) was performed to demonstrate the inhibitory effect of HMGB1 secretion by lipid raft disruption. It was found that release of HMGB1 blocked by nystatin as shown by NAC, a scavenger of ROS. Blot shown is representative of three experiments with similar results.
Reference
-
1. Fondevila C, Busuttil RW, Kupiec-Weglinski JW. Hepatic ischemia/reperfusion injury-a fresh look. Exp Mol Pathol. 2003. 74:86–93.2. Ock J, Cho H, Hong S, Kim IK, Suk K. Hypoxia as an initiator of neuroinflammation: microglial connections. Curr Neuropharmacol. 2005. 3:183–191.3. Akira S, Takeda K. Toll-like receptor signaling. Nat Rev Immunol. 2004. 4:499–511.4. Tsung A, Klune JR, Zhang X, Jeyabalan G, Cao Z, Peng X, et al. HMGB1 release induced by liver ischemia involves Toll-like receptor 4 dependent reactive oxygen species production and calcium-mediated signaling. J Exp Med. 2007. 204:2913–2923.5. Arumugam TV, Okun E, Tang SC, Thundyil J, Taylor SM, Woodruff TM. Toll-like receptors in ischemia-reperfusion injury. Shock. 2009. 32:4–16.6. Helms JB, Zurzolo C. Lipids as targeting signals: lipid rafts and intracellular trafficking. Traffic. 2004. 5:247–254.7. Hao M, Mukherjee S, Maxfield FR. Cholesterol depletion induces large scale domain segregation in living cell membranes. Proc Natl Acad Sci USA. 2001. 98:13072–13077.8. Kabouridis PS. Lipid rafts in T cell receptor signalling. Mol Membr Biol. 2006. 23:49–57.9. Hornef MW, Normark BH, Vandewalle A, Normark S. Intracellular recognition of lipopolysaccharide by toll-like receptor 4 in intestinal epithelial cells. J Exp Med. 2003. 198:1225–1235.10. Hirschfeld M, Ma Y, Weis JH, Vogel SN, Weis JJ. Cutting edge: repurification of lipopolysaccharide eliminates signaling through both human and murine tolllike receptor 2. J Immunol. 2000. 165:618–622.11. West MA, Billiar TR, Curran RD, Hyland BJ, Simmons RL. Evidence that rat Kupffer cells stimulate and inhibit hepatocyte protein synthesis in vitro by different mechanism. Gastroenterology. 1989. 96:1572–1582.12. Lai EC. Lipid rafts make for slippery platforms. J Cell Biol. 2003. 162:365–370.13. Triantafilou M, Triantafilou K. Lipopolysaccharide recognition: CD14, TLRs and the LPS-activation cluster. Trends Immunol. 2002. 23:301–304.14. Triantafilou M, Miyake K, Golenbock DT, Triantafilou K. Mediators of innate immune recognition of bacteria concentrate in lipid rafts and facilitate lipopolysaccharide-induced cell activation. J Cell Sci. 2002. 115:2603–2611.15. Marmor MD, Julius M. Role for lipid rafts in regulating interleukin-2 receptor signaling. Blood. 2001. 98:1489–1497.16. Miyake K. Endotoxin recognition molecules, Toll-like receptor 4-MD2. Semin Immunol. 2004. 16:11–16.17. Rothberg KG, Heuser JE, Donzell WC, Ying YS, Glenney JR, Anderson RG. Caveolin, a protein component of caveolae membrane coats. Cell. 1992. 68:673–682.18. Todd RF III, Petty HR. Beta 2 (CD11/CD18) integrins can serve as signaling partners for other leukocyte receptors. J Lab Clin Med. 1997. 129:492–498.19. Perera PY, Mayadas TN, Takeuchi O, Akira S, Zaks-Zilberman M, Goyert SM, et al. CD11b/CD18 acts in concert with CD14 and Toll-like receptor (TLR) 4 to elicit full lipopolysaccharide and taxol-inducible gene expression. J Immunol. 2001. 166:574–581.20. Jeyabalan G, Tsung A, Billiar TR. Linking proximal and downstream signaling events in hepatic ischemia/reperfusion injury. Biochem Soc Trans. 2006. 34:957–959.21. Tsung A, Sahai R, Tanaka H, Nakao A, Fink MP, Lotze MT, et al. The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion. J Exp Med. 2005. 201:1135–1143.22. Fan J, Li Y, Levy RM, Fan JJ, Hackam DJ, Vodovotz Y, et al. Hemorrhagic shock induces NAD(P)H oxidase activation in neutrophils: role of HMGB1-TLR4 signaling. J Immunol. 2007. 178:6573–6580.23. Miyake K. Roles for accessory molecules in microbial recognition by Toll-like receptors. J Endotoxin Res. 2006. 12:195–204.24. Szabo G, Dolganiuc A, Dai Q, Pruett SB. TLR4, ethanol, and lipid rafts: a new mechanism of ethanol action with implications for other receptor-mediated effects. J Immunol. 2007. 178:1243–1249.25. Dawson TL, Gores GL, Nieminen AL, Herman B, Lemasters JJ. Mitochondria as a source of reactive oxygen species during reductive stress in rat hepatocytes. Am J Physiol. 1993. 264:C961–C967.26. Chandel NS, McClintock DS, Feliciano CE, Wood TM, Melendez JA, Rodriguez AM, et al. Reactive oxygen species generated at mitochondrial complex III stabilize hypoxia-inducible factor-1 during hypoxia: a mechanism of O2 sensing. J Biol Chem. 2000. 275:25130–25138.27. Abdelrahman M, Mazzon E, Bauer M, Bauer J, Delbosc S, Cristol JP, et al. Inhibitors of NADPH oxidase reduce the organ injury in hemorrhagic shock. Shock. 2005. 23:107–114.28. Ulloa L, Messmer D. High-mobility group box 1 (HMGB1) protein: friend and foe. Cytokine Growth Factor Rev. 2006. 17:189–201.29. Wang H, Yang H, Tracey KJ. Extracellular role of HMGB1 in inflammation and sepsis. J Intern Med. 2004. 255:320–331.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Nucleic Acid Recognition and Signaling by Toll-like Receptor 9: Compartment-dependent Regulation
- Lipopolysaccharide: Basic Biochemistry, Intracellular Signaling, and Physiological Impacts in the Gut
- Toll-like receptor 4 antagonist and obesity associated kidney disease: Where should we go from here?
- Clinical Significance of Toll-Like Receptor and Toll-Like Receptor Blocker
- Targeting Microglial and Neuronal Toll-like Receptor 2 in Synucleinopathies
