The optimal model of reperfusion injury in vitro using H9c2 transformed cardiac myoblasts
- Affiliations
-
- 1Department of Pharmacology, University of Ulsan College of Medicine, Seoul 05505, Korea. kimyh@amc.seoul.kr
- 2Bio-Medical Institute of Technology, University of Ulsan, Seoul 05505, Korea.
- 3Stem Cell Immunomodulation Research Center, Asan Medical Center, University of Ulsan College of Medicine, Seoul 05505, Korea.
- 4Department of Medical Science, Asan Medical Institute of Convergence Science and Technology, Asan Medical Center, University of Ulsan College of Medicine, Seoul 05505, Korea.
- 5Asan Institute for Life Sciences, Asan Medical Center, Seoul 05505, Korea. cwwoo@amc.seoul.kr
- KMID: 2471039
- DOI: http://doi.org/10.4196/kjpp.2020.24.2.173
Abstract
- An in vitro model for ischemia/reperfusion injury has not been well-established. We hypothesized that this failure may be caused by serum deprivation, the use of glutamine-containing media, and absence of acidosis. Cell viability of H9c2 cells was significantly decreased by serum deprivation. In this condition, reperfusion damage was not observed even after simulating severe ischemia. However, when cells were cultured under 10% dialyzed FBS, cell viability was less affected compared to cells cultured under serum deprivation and reperfusion damage was observed after hypoxia for 24 h. Reperfusion damage after glucose or glutamine deprivation under hypoxia was not significantly different from that after hypoxia only. However, with both glucose and glutamine deprivation, reperfusion damage was significantly increased. After hypoxia with lactic acidosis, reperfusion damage was comparable with that after hypoxia with glucose and glutamine deprivation. Although high-passage H9c2 cells were more resistant to reperfusion damage than low-passage cells, reperfusion damage was observed especially after hypoxia and acidosis with glucose and glutamine deprivation. Cell death induced by reperfusion after hypoxia with acidosis was not prevented by apoptosis, autophagy, or necroptosis inhibitors, but significantly decreased by ferrostatin-1, a ferroptosis inhibitor, and deferoxamine, an iron chelator. These data suggested that in our SIR model, cell death due to reperfusion injury is likely to occur via ferroptosis, which is related with ischemia/reperfusion-induced cell death in vivo. In conclusion, we established an optimal reperfusion injury model, in which ferroptotic cell death occurred by hypoxia and acidosis with or without glucose/glutamine deprivation under 10% dialyzed FBS.
MeSH Terms
Figure
-
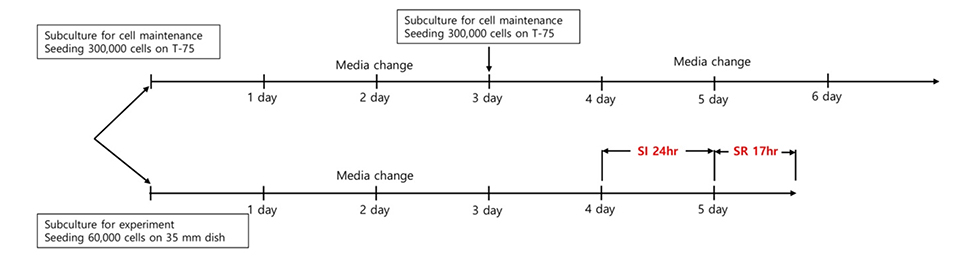
Fig. 1 Experimental schedule. SI stands for simulated ischemia and SR stands for simulated reperfusion. SI, simulated ischemia; SR, simulated reperfusion.
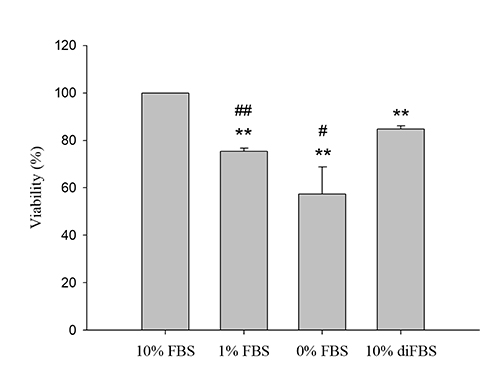
Fig. 2 Effect of fetal bovine serum (FBS) deprivation and 10% dialyzed FBS on H9c2 cell viability. To investigate the effect of serum deprivation and 10% dialyzed FBS on cell viability, MTT assay was performed. H9c2 cells were seeded in 6-well plates at 60,000 cells per well and cultured for four days. A media change was carried out once two days after seeding. After the medium was removed and cells were washed twice with PBS, DMEM was changed with FBS or dialyzed FBS at the concentration of the above experimental conditions. After culturing for 24 h, the medium was removed and 0.5 mg/ml Thiazolyl Blue Tetrazolium Blue (MTT) powder suspended in DMEM containing 10% FBS was treated. After 2 h, medium was removed, and cells were washed twice with PBS and subsequently treated with DMSO. The formazan solution was transferred to a 96-well plate and absorbance was measured at 562 nm. Data are expressed as means ± S.D. (n = 3) (**p < 0.01 vs. 10% FBS, and #p < 0.05 or ##p < 0.01 vs. 10% diFBS).
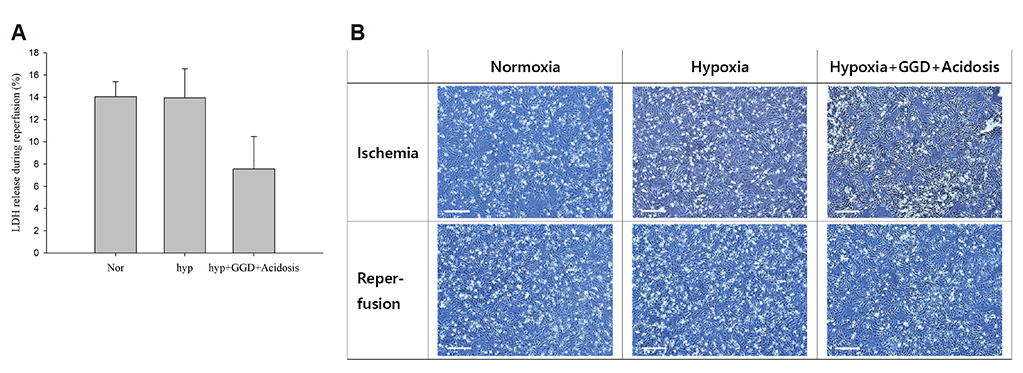
Fig. 3 Reperfusion damage under 1% serum condition. Ischemia/reperfusion simulation was performed under 1% fetal bovine serum (FBS) conditions. The control group was cultured in DMEM containing 1% FBS for 24 h under normoxia. The hypoxia group was cultured for 24 h under hypoxia (1% oxygen). To simulate extreme ischemia, cells were cultured under hypoxia, glucose/glutamine deprivation (GGD), and lactic acidosis (pH 6.4). Cells were cultured for 24 h in these conditions. After the ischemic period, the medium was changed with DMEM containing 1% FBS and incubated under normoxia. Seventeen hours after the media change, lactate dehydrogenase (LDH) release was measured. Cell photographs were taken as soon as ischemia was over and as soon as the reperfusion was over. Data are expressed as means ± S.D. (n = 3). Scale bar: 200 µm.

Fig. 4 Under 10% dialyzed fetal bovine serum (FBS), reperfusion damage was significantly increased when both glucose and glutamine were deprived. SnakeSkin wrinkle dialysis tubing was used to remove material ≤ 10 kDa in FBS, including nutrients such as glucose and glutamine. Dialyzed FBS was used for ischemia simulation with DMEM at a concentration of 10%. After culturing for 24 h under the specified conditions, cells were photographed. The medium was changed with DMEM containing 10% FBS. After 17 h incubation under normoxia, cells were photographed and lactate dehydrogenase (LDH) release was measured. Cell passage used in these experiments was below 20. Data are expressed as means ± S.D. (n = 3) (++p < 0.01 vs. control under normoxia, **p < 0.01 vs. control under hypoxia, #p < 0.05 vs. glucose/glutamine deprivation under hypoxia). Scale bar: 200 µm.

Fig. 5 Acidosis during hypoxia significantly induced reperfusion damage. To produce media with pH 6.4 under 5% CO2, 2.4 mM NaHCO3, obtained from the Henderson-Hasselbach equation, was added to DMEM (D5030). In the control group, 24 mM NaHCO3 was added to achieve pH 7.4. Therefore, we added 21.6 mM Na-lactate to match the osmolality of the media used to induce acidosis in the control group. As a result, we used the media, which is made for simulating lactic acidosis. Media pH was measured 7.4–7.5 in the control group and 6.4–6.5 in the acidosis group. Cell passage used in these experiments was below 20. Data are expressed as means ± S.D. (n = 3) (*p < 0.05 and **p < 0.01vs. control under hypoxia, #p < 0.05 vs. glucose/glutamine deprivation + acidosis under hypoxia). GGD, glucose/glutamine deprivation; N.S., not significant. Scale bar: 200 µm.
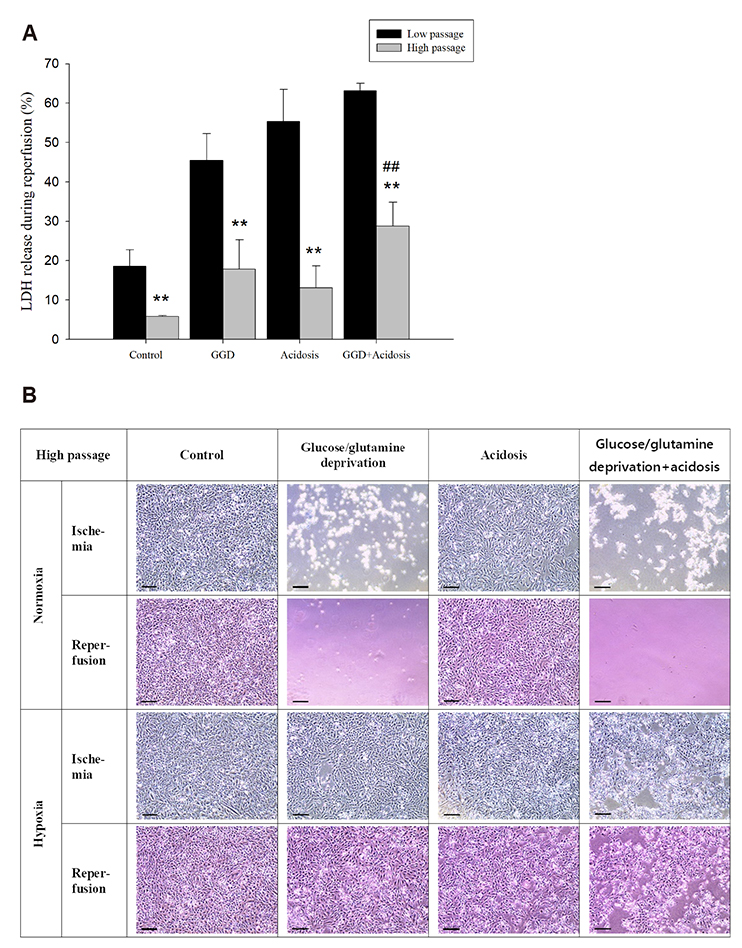
Fig. 6 High passage H9c2 cells obtained resistance to reperfusion damage. SIR was performed using high-passage H9c2 cells grown for 5 months or more. The passage used in the high passage data in (A) above was 25–29, and the passage of H9c2 used in the photograph of (B) was 27. Cells were maintained by subculturing twice weekly (Monday, Thursday). During subculturing, cell density did not exceed 90%, and media changed were also performed periodically (Wednesday, Saturday). There were no special events to mention while we maintain cells, such as extensive cell death or sudden changes in cell number. Data are expressed as means ± S.D. (high passage n = 4, low passage n = 3) (**p < 0.01 vs. low passage, ##p < 0.01vs. acidosis with hypoxia). LDH, lactate dehydrogenase; SIR, simulated ischemia/reperfusion; GGD, glucose/glutamine deprivation. Statistical analyses were conducted using an unpaired t-test. Scale bar: 200 µm.
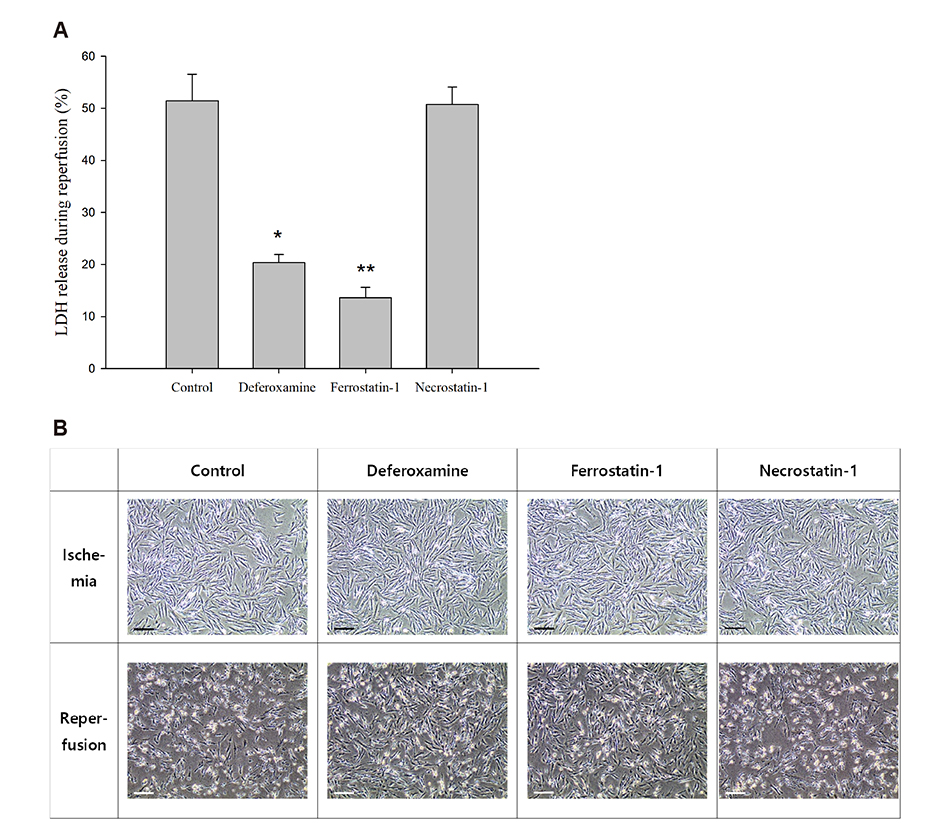
Fig. 7 Reperfusion induced iron-dependent cell death, which was likely ferroptosis. Before the media change to simulate the reperfusion, a media change was performed after adjusting the temperature and pH by incubating for 3–4 h in the incubator. At this time, 100 µM deferoxamine, 1 µM ferrostatin-1, and 20 µM necrostatin-1 were added to the media before incubation. Data are expressed as means ± S.D. (n = 3) (*p < 0.05 and **p < 0.01 vs. control). LDH, lactate dehydrogenase. Scale bar: 200 µm.
Reference
-
1. King LM, Opie LH. Glucose and glycogen utilisation in myocardial ischemia--changes in metabolism and consequences for the myocyte. Mol Cell Biochem. 1998; 180:3–26.2. Hearse DJ. Reperfusion of the ischemic myocardium. J Mol Cell Cardiol. 1977; 9:605–616.
Article3. Hausenloy DJ, Yellon DM. Myocardial ischemia-reperfusion injury: a neglected therapeutic target. J Clin Invest. 2013; 123:92–100.
Article4. Lee JW, Lee HK, Kim HW, Kim YH. Effects of pH, buffer system and lactate on the simulated ischemia-reperfusion injury of H9c2 cardiac myocytes. Korean J Physiol Pharmacol. 2007; 11:45–55.5. Yang GZ, Xue FS, Liu YY, Li HX, Liu Q, Liao X. Feasibility analysis of oxygen-glucose deprivation-nutrition resumption on H9c2 cells in vitro models of myocardial ischemia-reperfusion injury. Chin Med J (Engl). 2018; 131:2277–2286.6. Lee SB, Kim JJ, Kim TW, Kim BS, Lee MS, Yoo YD. Serum deprivation-induced reactive oxygen species production is mediated by Romo1. Apoptosis. 2010; 15:204–218.
Article7. Khogali SE, Harper AA, Lyall JA, Rennie MJ. Effects of L-glutamine on post-ischaemic cardiac function: protection and rescue. J Mol Cell Cardiol. 1998; 30:819–827.8. Gao M, Monian P, Quadri N, Ramasamy R, Jiang X. Glutaminolysis and transferrin regulate ferroptosis. Mol Cell. 2015; 59:298–308.
Article9. Steg PG, James SK, Gersh BJ. evidence-based recommendations, ensuring optimal patient management. Heart. 2013; 99:1156–1157.10. Bates ER, Jacobs AK. Time to treatment in patients with STEMI. N Engl J Med. 2013; 369:889–892.
Article11. Bond JM, Herman B, Lemasters JJ. Protection by acidotic pH against anoxia/reoxygenation injury to rat neonatal cardiac myocytes. Biochem Biophys Res Commun. 1991; 179:798–803.
Article12. Li L, Hao Y, Zhao Y, Wang H, Zhao X, Jiang Y, Gao F. Ferroptosis is associated with oxygen-glucose deprivation/reoxygenation-induced Sertoli cell death. Int J Mol Med. 2018; 41:3051–3062.
Article13. Li Y, Feng D, Wang Z, Zhao Y, Sun R, Tian D, Liu D, Zhang F, Ning S, Yao J, Tian X. Ischemia-induced ACSL4 activation contributes to ferroptosis-mediated tissue injury in intestinal ischemia/reperfusion. Cell Death Differ. 2019; 26:2284–2299.
Article14. Fang X, Wang H, Han D, Xie E, Yang X, Wei J, Gu S, Gao F, Zhu N, Yin X, Cheng Q, Zhang P, Dai W, Chen J, Yang F, Yang HT, Linkermann A, Gu W, Min J, Wang F. Ferroptosis as a target for protection against cardiomyopathy. Proc Natl Acad Sci U S A. 2019; 116:2672–2680.
Article15. Bonavita F, Stefanelli C, Giordano E, Columbaro M, Facchini A, Bonafè F, Caldarera CM, Guarnieri C. H9c2 cardiac myoblasts undergo apoptosis in a model of ischemia consisting of serum deprivation and hypoxia: inhibition by PMA. FEBS Lett. 2003; 536:85–91.
Article16. Kues WA, Anger M, Carnwath JW, Paul D, Motlik J, Niemann H. Cell cycle synchronization of porcine fetal fibroblasts: effects of serum deprivation and reversible cell cycle inhibitors. Biol Reprod. 2000; 62:412–419.17. Howard MK, Burke LC, Mailhos C, Pizzey A, Gilbert CS, Lawson WD, Collins MK, Thomas NS, Latchman DS. Cell cycle arrest of proliferating neuronal cells by serum deprivation can result in either apoptosis or differentiation. J Neurochem. 1993; 60:1783–1791.
Article18. Jain M, Cui L, Brenner DA, Wang B, Handy DE, Leopold JA, Loscalzo J, Apstein CS, Liao R. Increased myocardial dysfunction after ischemia-reperfusion in mice lacking glucose-6-phosphate dehydrogenase. Circulation. 2004; 109:898–903.
Article19. Le A, Lane AN, Hamaker M, Bose S, Gouw A, Barbi J, Tsukamoto T, Rojas CJ, Slusher BS, Zhang H, Zimmerman LJ, Liebler DC, Slebos RJ, Lorkiewicz PK, Higashi RM, Fan TW, Dang CV. Glucose-independent glutamine metabolism via TCA cycling for proliferation and survival in B cells. Cell Metab. 2012; 15:110–121.20. Fan J, Kamphorst JJ, Mathew R, Chung MK, White E, Shlomi T, Rabinowitz JD. Glutamine-driven oxidative phosphorylation is a major ATP source in transformed mammalian cells in both normoxia and hypoxia. Mol Syst Biol. 2013; 9:712.
Article21. DeBerardinis RJ, Mancuso A, Daikhin E, Nissim I, Yudkoff M, Wehrli S, Thompson CB. Beyond aerobic glycolysis: transformed cells can engage in glutamine metabolism that exceeds the requirement for protein and nucleotide synthesis. Proc Natl Acad Sci U S A. 2007; 104:19345–19350.
Article22. Son J, Lyssiotis CA, Ying H, Wang X, Hua S, Ligorio M, Perera RM, Ferrone CR, Mullarky E, Shyh-Chang N, Kang Y, Fleming JB, Bardeesy N, Asara JM, Haigis MC, DePinho RA, Cantley LC, Kimmelman AC. Glutamine supports pancreatic cancer growth through a KRAS-regulated metabolic pathway. Nature. 2013; 496:101–105.
Article23. Gao M, Yi J, Zhu J, Minikes AM, Monian P, Thompson CB, Jiang X. Role of Mitochondria in Ferroptosis. Mol Cell. 2019; 73:354–363.e3.
Article24. Myers RA. Hyperbaric oxygen therapy for trauma: crush injury, compartment syndrome, and other acute traumatic peripheral ischemias. Int Anesthesiol Clin. 2000; 38:139–151.
Article25. Kitakaze M, Weisfeldt ML, Marban E. Acidosis during early reperfusion prevents myocardial stunning in perfused ferret hearts. J Clin Invest. 1988; 82:920–927.
Article26. Inserte J, Barba I, Hernando V, Abellán A, Ruiz-Meana M, Rodríguez-Sinovas A, Garcia-Dorado D. Effect of acidic reperfusion on prolongation of intracellular acidosis and myocardial salvage. Cardiovasc Res. 2008; 77:782–790.
Article27. Khacho M, Tarabay M, Patten D, Khacho P, MacLaurin JG, Guadagno J, Bergeron R, Cregan SP, Harper ME, Park DS, Slack RS. Acidosis overrides oxygen deprivation to maintain mitochondrial function and cell survival. Nat Commun. 2014; 5:3550.
Article28. Williams RE, Zweier JL, Flaherty JT. Treatment with deferoxamine during ischemia improves functional and metabolic recovery and reduces reperfusion-induced oxygen radical generation in rabbit hearts. Circulation. 1991; 83:1006–1014.
Article29. Elihu N, Anandasbapathy S, Frishman WH. Chelation therapy in cardiovascular disease: ethylenediaminetetraacetic acid, deferoxamine, and dexrazoxane. J Clin Pharmacol. 1998; 38:101–105.
Article30. Xie Y, Hou W, Song X, Yu Y, Huang J, Sun X, Kang R, Tang D. Ferroptosis: process and function. Cell Death Differ. 2016; 23:369–379.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Effects of pH, Buffer System and Lactate on the Simulated Ischemia-reperfusion Injury of H9c2 Cardiac Myocytes
- The Different Expression Patterns of HSP22, a Late Embryogenesis Abundant-like Protein, in Hypertrophic H9C2 Cells Induced by NaCl and Angiotensin II
- Ischemic postconditioning protects cardiomyocytes against ischemia/reperfusion injury by inducing MIP2
- Studies on Differentiation of Cardiac Myoblast Induced by Co-culture with Isolated Neonatal Rat Cardiac Myocytes
- Benzoylaconine improves mitochondrial function in oxygenglucose deprivation and reperfusion-induced cardiomyocyte injury by activation of the AMPK/PGC-1 axis
