Suppression of Heme Oxygenase-1 by Prostaglandin E2-Protein Kinase A-A-Kinase Anchoring Protein Signaling Is Central for Augmented Cyclooxygenase-2 Expression in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages
- Affiliations
-
- 1Department of Internal Medicine, Eulji Hospital, Eulji University School of Medicine, Seoul, Korea. ksh1134@eulji.ac.kr
- 2Department of Bio-Medical Laboratory Science, College of Health Science, Eulji University, Suongnam, Korea.
- KMID: 2260815
- DOI: http://doi.org/10.4168/aair.2013.5.5.329
Abstract
- PURPOSE
Prostaglandin (PG) E2 is an immunomodulatory lipid mediator generated mainly via the cyclooxygenase-2 (COX-2) pathway from arachidonic acid at sites of infection and inflammation. A positive feedback loop of PGE2 on COX-2 expression is critical for homeostasis during toll-like receptor (TLR)-mediated inflammatory processes. The mechanism of PGE2-regulated COX-2 expression remains poorly understood. The low-molecular-weight stress protein heme oxygenase-1 (HO-1) contributes to the anti-inflammatory, anti-oxidant and anti-apoptotic response against environmental stress.
METHODS
We explored the involvement of HO-1 on PGE2 regulation of LPS-induced COX-2 expression in RAW 264.7 macrophages.
RESULTS
LPS-induced COX-2 expression in RAW 264.7 macrophages was enhanced by exogenous PGE2 or cyclic AMP (cAMP) analogue and was suppressed by a COX inhibitor (indomethacin), a protein kinase A (PKA) inhibitor (KT5720), and A kinase anchoring protein (AKAP) disruptors (Ht31 and RIAD). This result suggests that the stimulatory effects of endogenous and exogenous PGE2 on COX-2 expression are mediated by a cAMP-PKA-AKAP-dependent pathway. The induction of HO-1 was observed in LPS-stimulated RAW 264.7 macrophages. This induction was suppressed by exogenous PGE2 and enhanced by blockage of the endogenous PGE2 effect by the PKA inhibitor or AKAP disruptors. In addition, HO-1 induction by the HO activator copper protoporphyrin suppressed LPS-induced COX-2 expression, which was restored by the addition of exogenous PGE2. The induction of HO-1 inhibited LPS-induced NF-kappaB p-65 nuclear expression and translocation.
CONCLUSIONS
AKAP plays an important role in PGE2 regulation of COX-2 expression, and the suppression of HO-1 by PGE2-cAMP-PKA-AKAP signaling helps potentiate the LPS-induced COX-2 expression through a positive feedback loop in RAW 264.7 macrophages.
MeSH Terms
-
Arachidonic Acid
Copper
Cyclic AMP
Cyclic AMP-Dependent Protein Kinases
Cyclooxygenase 2
Dinoprostone
Heme
Heme Oxygenase-1
Homeostasis
Inflammation
Intracellular Signaling Peptides and Proteins
Macrophages
NF-kappa B
Phosphotransferases
Toll-Like Receptors
Arachidonic Acid
Copper
Cyclic AMP
Cyclic AMP-Dependent Protein Kinases
Cyclooxygenase 2
Dinoprostone
Heme
Heme Oxygenase-1
Intracellular Signaling Peptides and Proteins
NF-kappa B
Phosphotransferases
Toll-Like Receptors
Figure
-

Fig. 1 The cyclic AMP/protein kinase A/A-kinase anchoring protein (cAMP/PKA/AKAP) axis is responsible for prostaglandin E2 (PGE2)-potentiated lipopolysaccharide (LPS)-induced COX-2 expression in RAW 264.7 macrophages. (A) Cells were pretreated with the COX inhibitor indomethacin (Indo, 5 µM) for 30 min followed by PGE2 (1 µM) for 10 min and incubated with LPS (1 µg/mL) for another 24 h. (B) Cells were incubated with dibutyryl cAMP (1 µM) for 10 min followed by LPS for 24 h. (C) Cells were pretreated with PKA inhibitor KT5720 (1 µM) for 30 min followed by PGE2 for 10 min before incubation for 24 h with LPS. (D) Cells were pretreated for 20 min with the AKAP/PKA RII-specific disruptor peptide Ht31 (25 µM) or the AKAP/PKA RI-specific disruptor RIAD (25 µM) followed by LPS for 24 h. In all experiments, cell lysates (30 µg protein) were subjected to Western blot analysis of COX-2 and β-actin, upper. Results from one experiment of three are shown. Relative expression of COX-2 was determined by densitometric analysis of immunoblots from three different experiments, normalized for β-actin expression, and expressed as percent of LPS alone, lower. The data are the means±S.E. values of three independent experiments, each performed in duplicate. *P<0.05.

Fig. 2 Heme oxygenase-1 (HO-1) activation results in decreased LPS-induced COX-2 expression. Cells were pretreated with the HO activator copper protoporphyrin (CoPP, 10 µM) or the HO inhibitor zinc protoporphyrin (ZnPP, 20 µM) for 10 min followed by LPS (1 µg/mL) or vehicle for 6 h (left) and 24 h (right). Cell lysates at the indicated time points were subject to Western blot analysis of COX-2, HO-1, and β-actin. The blots are representative of three independently performed experiments.
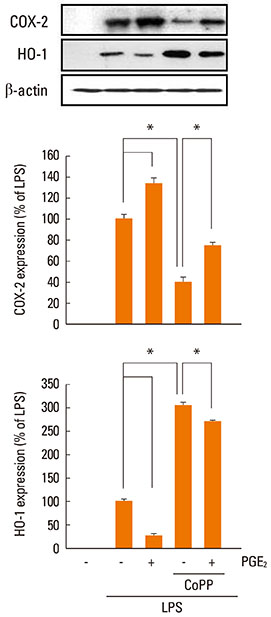
Fig. 3 Exogenous PGE2 suppresses LPS-induced HO-1 expression. Cells were pretreated with CoPP (10 µM) for 10 min followed by PGE2 (1 µM) for 10 min and then incubated with LPS (1 µg/mL) for 6 h. Lysates were subjected to Western blot analysis for COX-2, HO-1, and β-actin. Results from one experiment of three are shown. Relative expression levels of COX-2 and HO-1 were determined by densitometry analysis of immunoblots from three different experiments, normalized for β-actin expression, and expressed as percent of LPS alone. Data are the means±SEM of three independent experiments. *P<0.05.

Fig. 4 PKA/AKAP interaction is required for PGE2-suppressed effects on LPS-induced HO-1 expression. (A) Cells were pretreated with the PKA inhibitor KT5720 (1 µM) for 30 min followed by PGE2 for 10 min before incubation for 6 h with LPS. (B) Cells were pretreated with Ht31 (25 µM) or RIAD (25 µM) for 20 min and then incubated with LPS (1 µg/mL) for 6 h. In all experiments, lysates were subjected to Western blot analysis for HO-1 and β-actin. Results from one experiment of three are shown (upper). Relative expression of COX-2 and HO-1 were determined by densitometry analysis of immunoblots from three different experiments, normalized for β-actin expression, and expressed as percent of LPS alone (lower). Data represent the means±SEM of three separate experiments. *P<0.05.
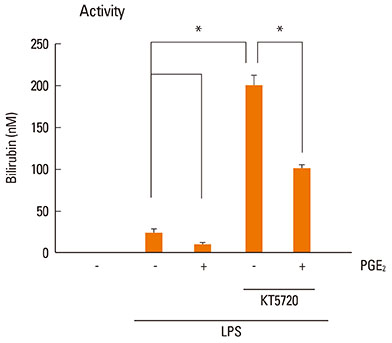
Fig. 5 Inhibition of PKA induces an increase in HO-1 activity. Cells were pretreated with the PKA inhibitor KT5720 (1 µM) for 20 min followed by PGE2 (1 µM) or vehicle for 10 min and then incubated with LPS for 6 h. Supernatants were harvested, and HO-1 activity was measured as described in the Materials and Methods. Data represent the means±SEM of three separate experiments. *P< 0.05.
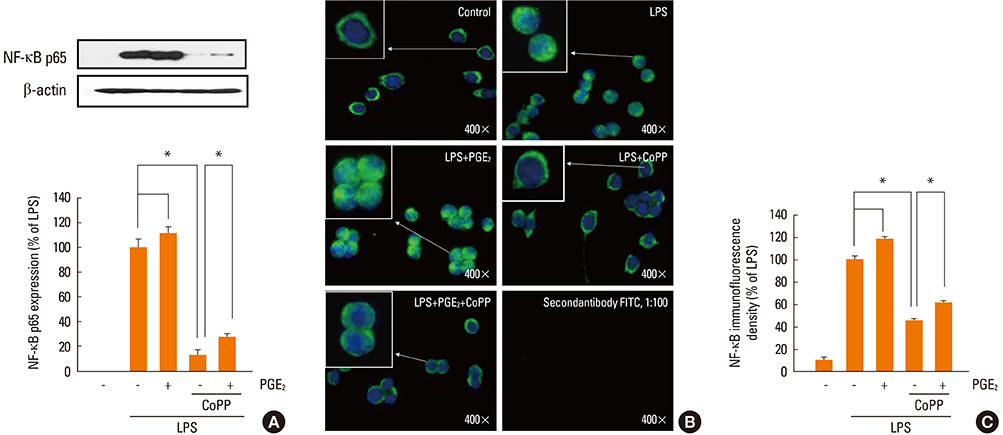
Fig. 6 Enhancement of HO-1 activity suppresses NF-κB signaling. (A) Cells were pretreated with vehicle control, or the HO activator CoPP (10 µM) for 10 min and then treated with PGE2 (1 µM) for 10 min before the addition of LPS (1 µg/mL). After 1 h, cells were harvested and the nuclear proteins were extracted as described in the Material and Methods. Immunoblotting of nuclear extracts was performed using anti-nuclear factor (NF)-κB p65. Results are representative of those from three experiments. *P<0.05. (B) Immunostaining for nuclear p65 localization in cells after 1 h exposure to: control, LPS only, LPS plus PGE2 (1 µM), LPS plus PGE2 plus CoPP, and LPS plus CoPP. Images depict nuclear translocation of NF-κB (arrows at green nuclei depict the translocation of NF-κB). (C) NF-κB immunofluorescence density determined by normalization with respect to translocation fluorescence. The nonfluorescent control cells were treated with second antibody only. Data are summarized and expressed as the means±SEM of three independent experiments. *P<0.05.
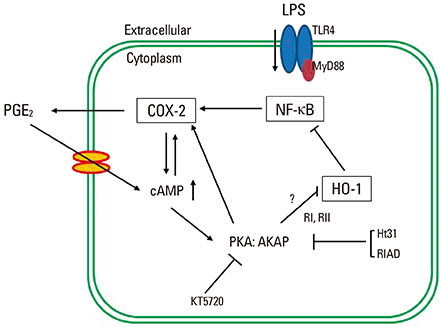
Fig. 7 Schematic representation of the proposed mechanism by which PGE2 signaling amplifies COX-2 expression through an autocrine manner in LPS-stimulated macrophages. Upon LPS binding to the toll-like receptor 4 (TLR4), the NF-κB signaling pathway becomes activated, leading to an increase in COX-2 expression, which in turn activates cAMP/PKA/AKAP through the production of PGE2. The PKA-AKAP complex blocks HO-1 activity, which results in enhanced translocation of NF-κB p65 to the nucleus.
Reference
-
1. Hwang D. Modulation of the expression of cyclooxygenase-2 by fatty acids mediated through toll-like receptor 4-derived signaling pathways. FASEB J. 2001; 15:2556–2564.2. Harris RE. Cyclooxygenase-2 (cox-2) blockade in the chemoprevention of cancers of the colon, breast, prostate, and lung. Inflammopharmacology. 2009; 17:55–67.3. Ren W, Hu L, Hua F, Jin J, Wang Y, Zhu L. Myeloid differentiation protein 2 silencing decreases LPS-induced cytokine production and TLR4/MyD88 pathway activity in alveolar macrophages. Immunol Lett. 2011; 141:94–101.4. Vancheri C, Mastruzzo C, Sortino MA, Crimi N. The lung as a privileged site for the beneficial actions of PGE2. Trends Immunol. 2004; 25:40–46.5. Serezani CH, Ballinger MN, Aronoff DM, Peters-Golden M. Cyclic AMP: master regulator of innate immune cell function. Am J Respir Cell Mol Biol. 2008; 39:127–132.6. Jarnaess E, Taskén K. Spatiotemporal control of cAMP signalling processes by anchored signalling complexes. Biochem Soc Trans. 2007; 35:931–937.7. Bos JL. Epac proteins: multi-purpose cAMP targets. Trends Biochem Sci. 2006; 31:680–686.8. Hinz B, Brune K, Pahl A. Prostaglandin E(2) upregulates cyclooxygenase-2 expression in lipopolysaccharide-stimulated RAW 264.7 macrophages. Biochem Biophys Res Commun. 2000; 272:744–748.9. Díaz-Muñoz MD, Osma-García IC, Fresno M, Iñiguez MA. Involvement of PGE2 and the cAMP signalling pathway in the upregulation of COX-2 and mPGES-1 expression in LPS-activated macrophages. Biochem J. 2012; 443:451–461.10. Hinz B, Brune K, Pahl A. Cyclooxygenase-2 expression in lipopolysaccharide-stimulated human monocytes is modulated by cyclic AMP, prostaglandin E(2), and nonsteroidal anti-inflammatory drugs. Biochem Biophys Res Commun. 2000; 278:790–796.11. Steinert D, Küper C, Bartels H, Beck FX, Neuhofer W. PGE2 potentiates tonicity-induced COX-2 expression in renal medullary cells in a positive feedback loop involving EP2-cAMP-PKA signaling. Am J Physiol Cell Physiol. 2009; 296:C75–C87.12. Paine A, Eiz-Vesper B, Blasczyk R, Immenschuh S. Signaling to heme oxygenase-1 and its anti-inflammatory therapeutic potential. Biochem Pharmacol. 2010; 80:1895–1903.13. Ryter SW, Alam J, Choi AM. Heme oxygenase-1/carbon monoxide: from basic science to therapeutic applications. Physiol Rev. 2006; 86:583–650.14. Chin BY, Otterbein LE. Carbon monoxide is a poison... to microbes! CO as a bactericidal molecule. Curr Opin Pharmacol. 2009; 9:490–500.15. Suh GY, Jin Y, Yi AK, Wang XM, Choi AM. CCAAT/enhancer-binding protein mediates carbon monoxide-induced suppression of cyclooxygenase-2. Am J Respir Cell Mol Biol. 2006; 35:220–226.16. Shih RH, Yang CM. Induction of heme oxygenase-1 attenuates lipopolysaccharide-induced cyclooxygenase-2 expression in mouse brain endothelial cells. J Neuroinflammation. 2010; 7:86.17. Leung PO, Wang SH, Lu SH, Chou WH, Shiau CY, Chou TC. Simvastatin inhibits pro-inflammatory mediators through induction of heme oxygenase-1 expression in lipopolysaccharide-stimulated RAW264.7 macrophages. Toxicol Lett. 2011; 207:159–166.18. Waltz P, Carchman EH, Young AC, Rao J, Rosengart MR, Kaczorowski D, Zuckerbraun BS. Lipopolysaccaride induces autophagic signaling in macrophages via a TLR4, heme oxygenase-1 dependent pathway. Autophagy. 2011; 7:315–320.19. Rhee SH, Hwang D. Murine TOLL-like receptor 4 confers lipopolysaccharide responsiveness as determined by activation of NF kappa B and expression of the inducible cyclooxygenase. J Biol Chem. 2000; 275:34035–34040.20. Chhikara M, Wang S, Kern SJ, Ferreyra GA, Barb JJ, Munson PJ, Danner RL. Carbon monoxide blocks lipopolysaccharide-induced gene expression by interfering with proximal TLR4 to NF-kappaB signal transduction in human monocytes. PLoS One. 2009; 4:e8139.21. Wall EA, Zavzavadjian JR, Chang MS, Randhawa B, Zhu X, Hsueh RC, Liu J, Driver A, Bao XR, Sternweis PC, Simon MI, Fraser ID. Suppression of LPS-induced TNF-alpha production in macrophages by cAMP is mediated by PKA-AKAP95-p105. Sci Signal. 2009; 2:ra28.22. Kim SH, Serezani CH, Okunishi K, Zaslona Z, Aronoff DM, Peters-Golden M. Distinct protein kinase A anchoring proteins direct prostaglandin E2 modulation of Toll-like receptor signaling in alveolar macrophages. J Biol Chem. 2011; 286:8875–8883.23. Olszanecki R, Kurnyta M, Biedroń R, Chorobik P, Bereta M, Marcinkiewicz J. The role of heme oxygenase-1 in down regulation of PGE2 production by taurine chloramine and taurine bromamine in J774.2 macrophages. Amino Acids. 2008; 35:359–364.24. Liu XH, Pan LL, Yang HB, Gong QH, Zhu YZ. Leonurine attenuates lipopolysaccharide-induced inflammatory responses in human endothelial cells: involvement of reactive oxygen species and NF-kappaB pathways. Eur J Pharmacol. 2012; 680:108–114.25. Immenschuh S, Kietzmann T, Hinke V, Wiederhold M, Katz N, Muller-Eberhard U. The rat heme oxygenase-1 gene is transcriptionally induced via the protein kinase A signaling pathway in rat hepatocyte cultures. Mol Pharmacol. 1998; 53:483–491.26. Kim HJ, Tsoy I, Park MK, Lee YS, Lee JH, Seo HG, Chang KC. Iron released by sodium nitroprusside contributes to heme oxygenase-1 induction via the cAMP-protein kinase A-mitogen-activated protein kinase pathway in RAW 264.7 cells. Mol Pharmacol. 2006; 69:1633–1640.27. Park MK, Kang YJ, Ha YM, Jeong JJ, Kim HJ, Seo HG, Lee JH, Chang KC. EP2 receptor activation by prostaglandin E2 leads to induction of HO-1 via PKA and PI3K pathways in C6 cells. Biochem Biophys Res Commun. 2009; 379:1043–1047.28. Nakao S, Ogata Y, Shimizu-Sasaki E, Yamazaki M, Furuyama S, Sugiya H. Activation of NFkappaB is necessary for IL-1beta-induced cyclooxygenase-2 (COX-2) expression in human gingival fibroblasts. Mol Cell Biochem. 2000; 209:113–118.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Anti-inflammatory effects of proanthocyanidin-rich red rice extract via suppression of MAPK, AP-1 and NF-κB pathways in Raw 264.7 macrophages
- TI-I-174, a Synthetic Chalcone Derivative, Suppresses Nitric Oxide Production in Murine Macrophages via Heme Oxygenase-1 Induction and Inhibition of AP-1
- Anti-inflammatory effect of methanol extract from Erigeron Canadensis L. may be involved with upregulation of heme oxygenase-1 expression and suppression of NFkappaB and MAPKs activation in macrophages
- Pulegone Exhibits Anti-inflammatory Activities through the Regulation of NF-κB and Nrf-2 Signaling Pathways in LPS-stimulated RAW 264.7 cells
- Black soybean anthocyanins attenuate inflammatory responses by suppressing reactive oxygen species production and mitogen activated protein kinases signaling in lipopolysaccharide-stimulated macrophages
