Functional Role of Parkin against Oxidative Stress in Neural Cells
- Affiliations
-
- 1Department of Biomedical Sciences, Korea University College of Medicine, Seoul, Korea. geumd@korea.ac.kr
- 2Department of Emergency Medical Technology, Kyungil University College of Nursing and Public Health, Gyeongsan, Korea. yhkim01@kiu.ac.kr
- KMID: 2169353
- DOI: http://doi.org/10.3803/EnM.2014.29.1.62
Abstract
- BACKGROUND
Parkinson disease (PD) is caused by selective cell death of dopaminergic neurons in the substantia nigra. An early onset form of PD, autosomal recessive juvenile parkinsonism has been associated with a mutation in the parkin gene. The function of parkin is known to remove misfolding proteins and protect cell death. We aimed to investigate the role of parkin against oxidative stress in neuronal cells.
METHODS
Parkin knockout embryonic stem cells (PKO ES cells) were differentiated into neurons by adherent monolayer culture method. Oxidative stress was induced by the treatment of 1-methyl-4-phenylpyridinium (MPP+) in neurons derived from wild type and PKO ES cells, and cell viability was examined by MTT assay. After exposure to MPP+, Tuj1-positive cell population was compared between PKO and wild type cells by fluorescence activated cell sorter (FACS) analysis. The activated caspase3 protein level was also measured by Western blot analysis, FACS and immunocytochemistry.
RESULTS
There was no difference in the efficiency of neuronal differentiation between wild type and PKO ES cells. After exposure to MPP+, no significant differences were found in cell viability and Tuj1-positive cell population between the two groups determined by MTT assay and FACS analysis, respectively. The activated caspase3 protein levels examined by Western blot analysis, FACS and immunocytochemistry were not changed in PKO cells compared with those of wild type cells after MPP+ treatment.
CONCLUSION
These results suggest that PKO neuronal cells including dopaminergic neurons are not sensitive to caspase3-dependent cell death pathway during the response against MPP+-induced oxidative stress.
MeSH Terms
Figure
-
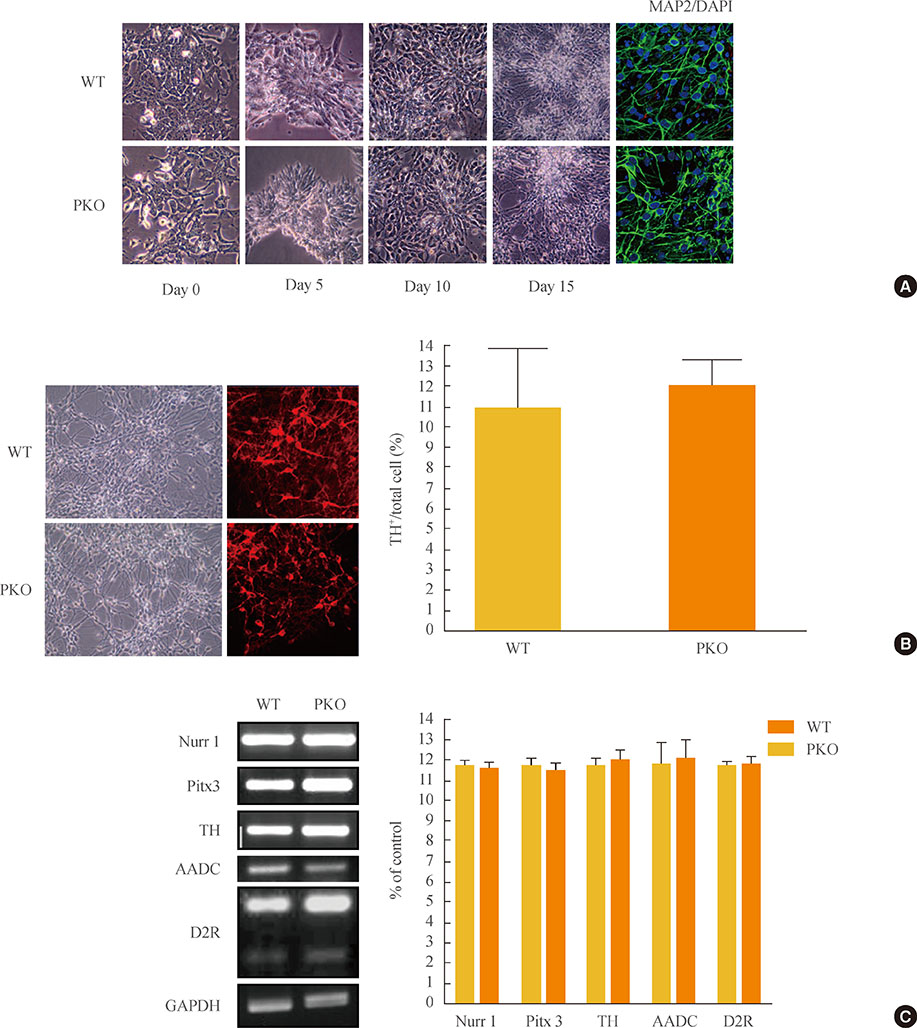
Fig. 1 Induction of dopaminergic neurons from wild-type (WT) and parkin knockout embryonic stem (PKO ES) cells by the adherent monolayer culture method. (A) Induction of neural cells from WT and PKO ES cells by the adherent monolayer method. Representative images were taken 0, 5, 10, and 15 days after differentiation. Immunocytochemistry shows MAP2-positive mature neural cells, and 6-diamidino-2-phenylindole (DAPI) was used for nuclear staining. (B) Expression of tyrosine hydroxylase (TH) 15 days after differentiation. TH+ neurons were analyzed by immunocytochemistry with an anti-TH antibody 15 days after differentiation. The 10% to 12% of cells were identified as TH-positive cells. (C) Reverse-transcriptional polymerase chain reaction analysis of dopaminergic neuron markers, such as Nurr1, TH, Pitx3, aromatic L-amino acid decarboxylase (AADC), and dopamine receptor 2 (D2R) at 15 days after differentiation. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control. Data are expressed as mean±SEM (n=3 per group). Statistical analysis was performed by Student t test. No significant differences in dopaminergic neuron marker expression were observed between WT and PKO cells.

Fig. 2 Cell viability analysis after treatment with 1-methyl-4-phenylpyridinium (MPP+) by MTT assay. MPP+ was treated at various concentrations (0, 100, 500, 750, 1,000, 1,250, and 1,500 µM) for 24 hours, at 15 days after differentiation. MTT assay was performed for viability of differentiated cells. Data are expressed as mean±SEM (n=6 per group). Statistical analysis was performed by two-way analysis of variance. No significant differences in the MPP+ susceptibility were observed between wild-type (WT) and parkin knockout (PKO) cells.
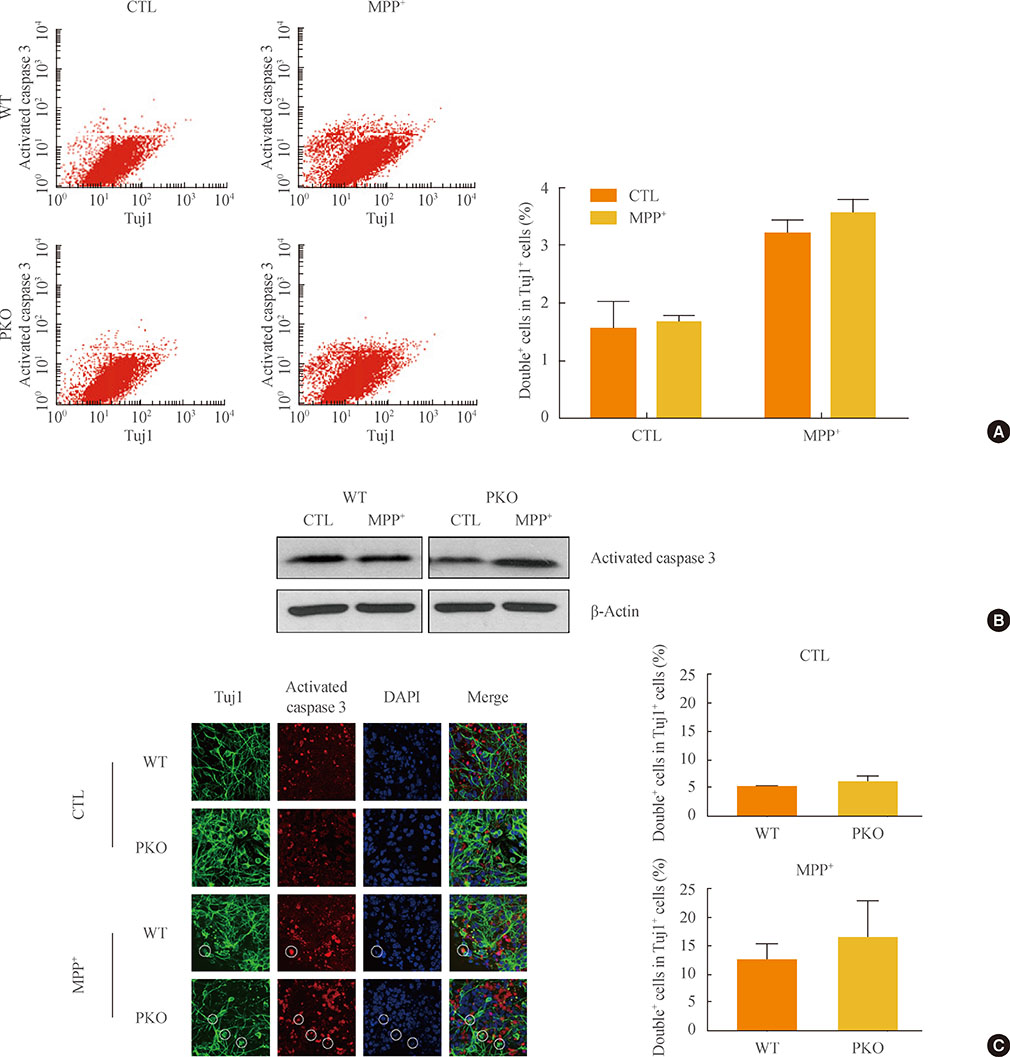
Fig. 3 Activity of caspase 3 in 1-methyl-4-phenylpyridinium (MPP+) treated wild-type (WT) and parkin knockout (PKO) neuronal cells. (A) Fluorescence-activated cell sorting analysis of WT and PKO cells treated with 1 mM MPP+ for 24 hours with anti-Tuj1 and antiactivated caspase 3 antibodies. Double+ cells represent colabeling with Tuj1 and activated caspase 3. The right panel shows the percentage of double+ cells per Tuj1 positive cells. (B) Western blot analysis of caspase 3 activity in WT and PKO cells. Cells were harvested 24 hours after 1 mM MPP+ treatment and Western blot analysis was performed with an activated caspase 3 antibody. β-Actin was used as an internal control. (C) Immunocytochemistry of WT and PKO cells treated with 1 mM MPP+ for 24 hours with anti-Tuj1 (green) and activated caspase 3 (red) antibody (scale bar, 20 µm). While MPP+ treatment causes a significant increase in caspase 3 activity, no significant differences in caspase 3 activity are observed between WT and PKO neural cells. Data are expressed as the mean±SEM (n=3 per group). Statistical analysis was performed by Student t test. CTL, control without MPP+ treatment; DAPI, 4, 6-diamidino-2-phenylindole.
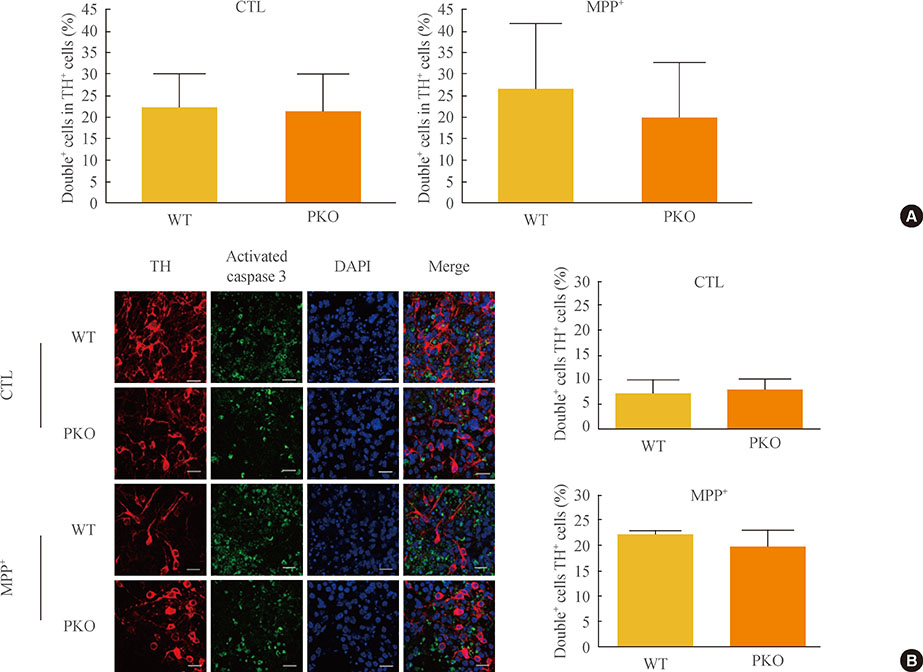
Fig. 4 Activity of caspase 3 in 1-methyl-4-phenylpyridinium (MPP+) treated wild-type (WT) and parkin knockout (PKO) dopaminergic neurons. (A) Fluorescence-activated cell sorting analysis of WT and PKO cells treated with 1 mM MPP+ for 24 hours with tyrosine hydroxylase (TH) and activated caspase 3 antibodies. Double+ cells represent colabeling with TH and activated caspase 3 antibodies. Graphs show the percentage of double positive cells per TH-positive cells. (B) Immunocytochemistry of WT and PKO cells treated with 1 mM MPP+ for 24 hours with TH (red) and activated caspase 3 (green) antibodies (scale bar, 20 µm). No significant differences in caspase 3 activity were observed between WT and PKO dopaminergic neurons. Data are expressed as mean±SEM (n=3 per group). Statistical analysis was performed by Student t test. CTL, control without MPP+ treatment; DAPI, 4, 6-diamidino-2-phenylindole.
Reference
-
1. Bossy-Wetzel E, Schwarzenbacher R, Lipton SA. Molecular pathways to neurodegeneration. Nat Med. 2004; 10:Suppl. S2–S9.2. Taylor JP, Hardy J, Fischbeck KH. Toxic proteins in neurodegenerative disease. Science. 2002; 296:1991–1995.3. Alam ZI, Jenner A, Daniel SE, Lees AJ, Cairns N, Marsden CD, Jenner P, Halliwell B. Oxidative DNA damage in the parkinsonian brain: an apparent selective increase in 8-hydroxyguanine levels in substantia nigra. J Neurochem. 1997; 69:1196–1203.4. Arlt S, Beisiegel U, Kontush A. Lipid peroxidation in neurodegeneration: new insights into Alzheimer's disease. Curr Opin Lipidol. 2002; 13:289–294.5. Gabbita SP, Lovell MA, Markesbery WR. Increased nuclear DNA oxidation in the brain in Alzheimer's disease. J Neurochem. 1998; 71:2034–2040.6. Pedersen WA, Fu W, Keller JN, Markesbery WR, Appel S, Smith RG, Kasarskis E, Mattson MP. Protein modification by the lipid peroxidation product 4-hydroxynonenal in the spinal cords of amyotrophic lateral sclerosis patients. Ann Neurol. 1998; 44:819–824.7. Finkel T, Holbrook NJ. Oxidants, oxidative stress and the biology of ageing. Nature. 2000; 408:239–247.8. Shimura H, Hattori N, Kubo S, Mizuno Y, Asakawa S, Minoshima S, Shimizu N, Iwai K, Chiba T, Tanaka K, Suzuki T. Familial Parkinson disease gene product, parkin, is a ubiquitin-protein ligase. Nat Genet. 2000; 25:302–305.9. Palacino JJ, Sagi D, Goldberg MS, Krauss S, Motz C, Wacker M, Klose J, Shen J. Mitochondrial dysfunction and oxidative damage in parkin-deficient mice. J Biol Chem. 2004; 279:18614–18622.10. Clark IE, Dodson MW, Jiang C, Cao JH, Huh JR, Seol JH, Yoo SJ, Hay BA, Guo M. Drosophila pink1 is required for mitochondrial function and interacts genetically with parkin. Nature. 2006; 441:1162–1166.11. Park J, Lee SB, Lee S, Kim Y, Song S, Kim S, Bae E, Kim J, Shong M, Kim JM, Chung J. Mitochondrial dysfunction in Drosophila PINK1 mutants is complemented by parkin. Nature. 2006; 441:1157–1161.12. Dodson MW, Guo M. Pink1, Parkin, DJ-1 and mitochondrial dysfunction in Parkinson's disease. Curr Opin Neurobiol. 2007; 17:331–337.13. Ying QL, Stavridis M, Griffiths D, Li M, Smith A. Conversion of embryonic stem cells into neuroectodermal precursors in adherent monoculture. Nat Biotechnol. 2003; 21:183–186.14. Darios F, Corti O, Lucking CB, Hampe C, Muriel MP, Abbas N, Gu WJ, Hirsch EC, Rooney T, Ruberg M, Brice A. Parkin prevents mitochondrial swelling and cytochrome c release in mitochondria-dependent cell death. Hum Mol Genet. 2003; 12:517–526.15. Goldberg MS, Fleming SM, Palacino JJ, Cepeda C, Lam HA, Bhatnagar A, Meloni EG, Wu N, Ackerson LC, Klapstein GJ, Gajendiran M, Roth BL, Chesselet MF, Maidment NT, Levine MS, Shen J. Parkin-deficient mice exhibit nigrostriatal deficits but not loss of dopaminergic neurons. J Biol Chem. 2003; 278:43628–43635.16. Langston JW, Ballard P, Tetrud JW, Irwin I. Chronic Parkinsonism in humans due to a product of meperidine-analog synthesis. Science. 1983; 219:979–980.17. Schapira AH, Cooper JM, Dexter D, Jenner P, Clark JB, Marsden CD. Mitochondrial complex I deficiency in Parkinson's disease. Lancet. 1989; 1:1269.18. Dipasquale B, Marini AM, Youle RJ. Apoptosis and DNA degradation induced by 1-methyl-4-phenylpyridinium in neurons. Biochem Biophys Res Commun. 1991; 181:1442–1448.19. Du Y, Dodel RC, Bales KR, Jemmerson R, Hamilton-Byrd E, Paul SM. Involvement of a caspase-3-like cysteine protease in 1-methyl-4-phenylpyridinium-mediated apoptosis of cultured cerebellar granule neurons. J Neurochem. 1997; 69:1382–1388.20. Hartley A, Stone JM, Heron C, Cooper JM, Schapira AH. Complex I inhibitors induce dose-dependent apoptosis in PC12 cells: relevance to Parkinson's disease. J Neurochem. 1994; 63:1987–1990.21. Itano Y, Nomura Y. 1-methyl-4-phenyl-pyridinium ion (MPP+) causes DNA fragmentation and increases the Bcl-2 expression in human neuroblastoma, SH-SY5Y cells, through different mechanisms. Brain Res. 1995; 704:240–245.22. Thomas B, von Coelln R, Mandir AS, Trinkaus DB, Farah MH, Leong Lim K, Calingasan NY, Flint Beal M, Dawson VL, Dawson TM. MPTP and DSP-4 susceptibility of substantia nigra and locus coeruleus catecholaminergic neurons in mice is independent of parkin activity. Neurobiol Dis. 2007; 26:312–322.23. Fallon L, Belanger CM, Corera AT, Kontogiannea M, Regan-Klapisz E, Moreau F, Voortman J, Haber M, Rouleau G, Thorarinsdottir T, Brice A, van Bergen En Henegouwen PM, Fon EA. A regulated interaction with the UIM protein Eps15 implicates parkin in EGF receptor trafficking and PI(3)K-Akt signalling. Nat Cell Biol. 2006; 8:834–842.24. Henn IH, Bouman L, Schlehe JS, Schlierf A, Schramm JE, Wegener E, Nakaso K, Culmsee C, Berninger B, Krappmann D, Tatzelt J, Winklhofer KF. Parkin mediates neuroprotection through activation of IkappaB kinase/nuclear factor-kappaB signaling. J Neurosci. 2007; 27:1868–1878.25. Jiang H, Jiang Q, Feng J. Parkin increases dopamine uptake by enhancing the cell surface expression of dopamine transporter. J Biol Chem. 2004; 279:54380–54386.26. Jiang H, Ren Y, Zhao J, Feng J. Parkin protects human dopaminergic neuroblastoma cells against dopamine-induced apoptosis. Hum Mol Genet. 2004; 13:1745–1754.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Epilepsy and Oxidative Stress
- Reactive Oxygen Species: Role in Pathophysiology, and Mechanism of Endogenous and Dietary Antioxidants during Oxidative Stress
- The Role of Oxidative Stress in the Pathogenesis of Asthma
- The role of oxidative stress and hypoxia in renal disease
- Effect of Hydrogen Peroxide-induced Oxidative Stress on the Senescence of Trabecular Meshwork Cells
