Arsenic trioxide induces depolymerization of microtubules in an acute promyelocytic leukemia cell line
- Affiliations
-
- 1Division of Hematology and Oncology, Department of Internal Medicine, Ulsan University Hospital, University of Ulsan College of Medicine, Ulsan, Korea. yjmin@uuh.ulsan.kr
- 2Biomedical Research Center, Ulsan University Hospital, University of Ulsan College of Medicine, Ulsan, Korea.
- 3Division of Hematology-Oncology, Soonchunhyang University College of Medicine, Seoul, Korea.
- KMID: 2083603
- DOI: http://doi.org/10.5045/kjh.2012.47.2.105
Abstract
- BACKGROUND
Arsenic trioxide (As2O3) is a well-known and effective treatment that can result in clinical remission for patients diagnosed with acute promyelocytic leukemia (APL). The biologic efficacy of As2O3 in APL and solid tumor cells has been explained through its actions on anti-proliferation, anti-angiogenesis, and apoptotic signaling pathways. We theorize that As2O3 activates a pathway that disrupts microtubule dynamics forming abnormal, nonfunctioning mitotic spindles, thus preventing cellular division. In this study, we investigated how As2O3 induces apoptosis by causing microtubule dysfunction.
METHODS
Cultured NB4 cells were treated with As2O3, paclitaxel, and vincristine. Flow cytometric analysis was then performed. An MTT assay was used to determine drug-mediated cytotoxicity. For tubulin polymerization assay, each polymerized or soluble tubulin was measured. Microtubule assembly-disassembly was measured using a tubulin polymerization kit. Cellular microtubules were also observed with fluorescence microscopy.
RESULTS
As2O3 treatment disrupted tubulin assembly resulting in dysfunctional microtubules that cause death in APL cells. As2O3 markedly enhanced the amount of depolymerized microtubules. The number of microtubule posttranslational modifications on an individual tubulin decreased with As2O3 concentration. Immunocytochemistry revealed changes in the cellular microtubule network and formation of polymerized microtubules in As2O3-treated cells.
CONCLUSION
The microtubules alterations found with As2O3 treatment suggest that As2O3 increases the depolymerized forms of tubulin in cells and that this is potentially due to arsenite's negative effects on spindle dynamics.
MeSH Terms
-
Antimitotic Agents
Apoptosis
Arsenic
Arsenicals
Cell Line
Fluorescence
Humans
Immunohistochemistry
Leukemia, Promyelocytic, Acute
Microtubules
Oxides
Paclitaxel
Polymerization
Polymers
Protein Processing, Post-Translational
Tubulin
Vincristine
Antimitotic Agents
Arsenic
Arsenicals
Oxides
Paclitaxel
Polymers
Tubulin
Vincristine
Figure
-
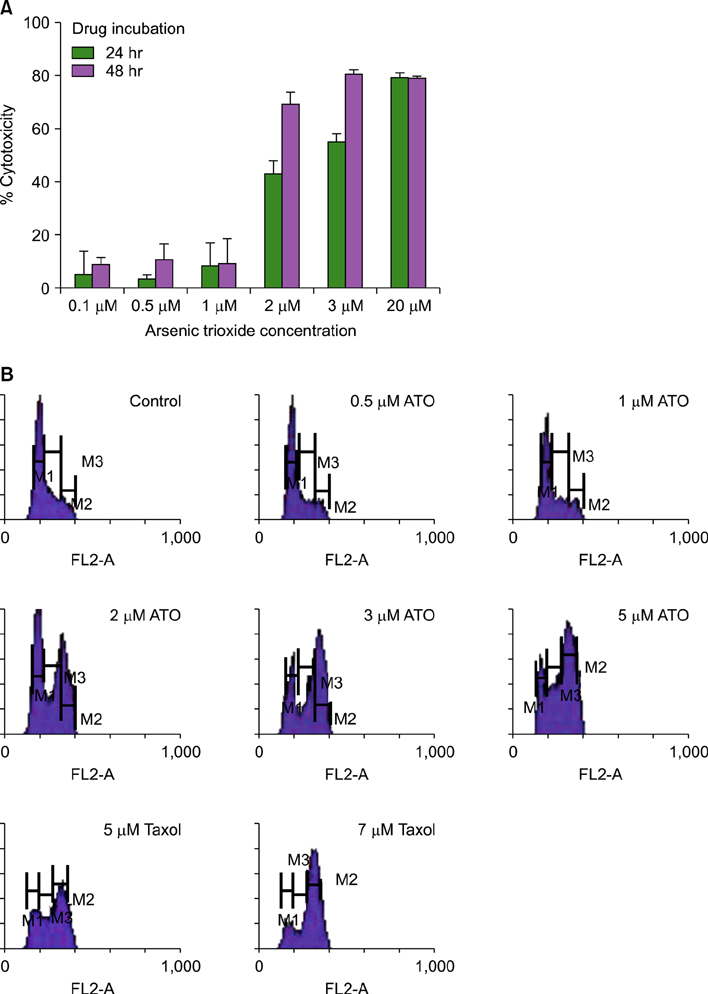
Fig. 1 Induction of cytotoxicity and cell cycle arrest by As2O3. (A) To show the inhibitory effects of As2O3 on the cell proliferation, exponentially growing cells were treated with the indicated concentrations of As2O3 for 24 hour and 48 hours, and cell proliferation was assessed by using the MTS reagent (see Materials and Methods). The cytotoxic effect of As2O3 was increased in a concentration- and/or time-dependent manner. These data represent the mean±SD of independent experiments. (B) To observe the apoptotic effects of As2O3, NB4 cells were treated with either a vehicle (control) or each concentration of As2O3 and paclitaxel for 24 hours. PI-stained cells were analyzed by flow cytometry as described in the Materials and Methods section, and the percentage of cells in each phase of the cell cycle was analyzed using flow cytometry (Table 1). Flow cytometry analysis revealed a significant increased apoptotic effect that resulted in a concentration-dependent accumulation of NB4 cells in the G2/M phase on the As2O3-treated cells.

Fig. 2 Alterations in tubulin polymerization induced by As2O3. (A) To demonstrate that the fraction of depolymerized tubulin is increased after treatment with As2O3, NB4 cells were treated with drugs or without drugs as a control for 6 hours at the indicated concentrations. Cell lysates were separated into polymerized (P) or soluble (S) fractions. Aliquots of equal volume were separated on an SDS-PAGE gel, and evaluated by western blotting with an anti α-tubulin antibody. Compared to the baseline proportion of α-tubulin in the soluble fraction, the depolymerized proportion observed after treatment of As2O3 was approximately 52-72% and the tubulin shift to the "S" fraction was noted. (B) To show the effects of a combination of drugs, NB4 cells were treated with 5 µM As2O3, 10 µM As2O3 and 40 nM paclitaxel at alone or in combination, and treated for 6 hours. Cell lysates were separated into "P" or "S" fractions as described above. Aliquots of equal volume were loaded onto SDS-PAGE gels, and the blots probed with antibody against α-tubulin. The results indicate that As2O3 and paclitaxel did not synergistically influence microtubule assembly. The intensity of each band was quantified by densitometry and the percentage of soluble tubulin (%S) was calculated by multiplying the fraction of tubulin in the soluble fraction [S/(S+P)] by 100 for each "S-P" pair.
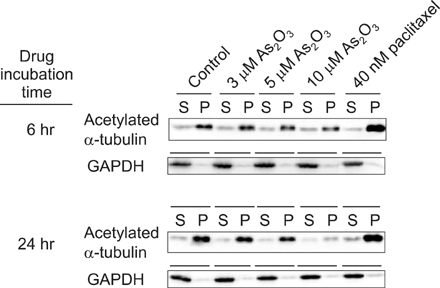
Fig. 3 Effects of As2O3 on microtubule stabilization. NB4 cells were treated with drugs or no drugs as a control, for 6 hours and 24 hours at the indicated concentration. Lysates were separated into polymerized (P) or soluble (S) fractions. The blots probed with anti-acetylated α-tubulin. The amount of acetylated α-tubulin in the P fraction of paclitaxel treatment was increased, compared with that of the control, but the proportion in P fraction of As2O3 treatment was decreased in a concentration-dependent manner. This result indicated that As2O3 interfered with important modifications necessary for the stability of microtubules. The intensity of each band was quantified by densitometry and the blots were stripped and reprobed with GAPDH-specific antibody as a loading control.
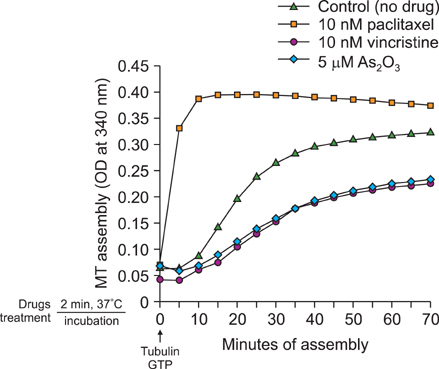
Fig. 4 Effects of As2O3 on microtubule polymerization in vitro. Purified tubulin from bovine brain tissue (Cytoskeleton) was incubated at 37℃ in reaction mixtures containing 1 mM GTP, 10 nM paclitaxel (Taxol), 10 nM vincristine, 5 µM As2O3 and the mock-treated solution as a control. Tubulin polymerization was determined by measuring absorbance at 340 nm. This revealed that As2O3 directly influences tubulin and induces tubulin depolymerization.
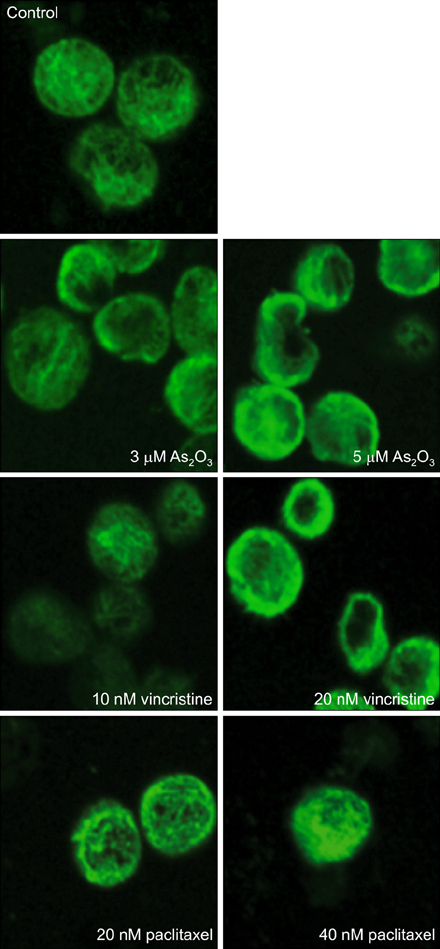
Fig. 5 Effects of As2O3 on the organization of cellular microtubule network. NB4 cells were treated with 5 µM As2O3, 40 nM paclitaxel, and 20 nM vincristine. Mock-treated cells were used as a control. After a 6-hour incubation, cells were harvested and fixed with formaldehyde. Cells were incubated with monoclonal anti-α-tubulin antibody at room temperature for 30 minutes. After incubation with FITC-conjugated secondary antibody, the cellular microtubules were imaged using an Olympus IX-81 fluorescence microscope. The normal organization of microtubule network was seen in control cells, increased density of polymerized microtubules were found in paclitaxel-treated cells, and a degraded microtubule network in cytoplasm was observed in As2O3-and vicristine-treated cells.
Reference
-
1. Kamimura T, Miyamoto T, Harada M, Akashi K. Advances in therapies for acute promyelocytic leukemia. Cancer Sci. 2011. 102:1929–1937.
Article2. Miller WH Jr, Schipper HM, Lee JS, Singer J, Waxman S. Mechanisms of action of arsenic trioxide. Cancer Res. 2002. 62:3893–3903.3. Fojo T, Menefee M. Mechanisms of multidrug resistance: the potential role of microtubule-stabilizing agents. Ann Oncol. 2007. 18:Suppl 5. v3–v8.
Article4. Barlow SB, Gonzalez-Garay ML, Cabral F. Paclitaxel-dependent mutants have severely reduced microtubule assembly and reduced tubulin synthesis. J Cell Sci. 2002. 115:3469–3478.
Article5. Kavallaris M, Tait AS, Walsh BJ, et al. Multiple microtubule alterations are associated with Vinca alkaloid resistance in human leukemia cells. Cancer Res. 2001. 61:5803–5809.6. Jordan MA, Wilson L. Microtubules as a target for anticancer drugs. Nat Rev Cancer. 2004. 4:253–265.
Article7. Huang SC, Lee TC. Arsenite inhibits mitotic division and perturbs spindle dynamics in HeLa S3 cells. Carcinogenesis. 1998. 19:889–896.
Article8. Ling YH, Jiang JD, Holland JF, Perez-Soler R. Arsenic trioxide produces polymerization of microtubules and mitotic arrest before apoptosis in human tumor cell lines. Mol Pharmacol. 2002. 62:529–538.
Article9. Li YM, Broome JD. Arsenic targets tubulins to induce apoptosis in myeloid leukemia cells. Cancer Res. 1999. 59:776–780.10. Yan J, Xu YH. Tributyrin inhibits human gastric cancer SGC-7901 cell growth by inducing apoptosis and DNA synthesis arrest. World J Gastroenterol. 2003. 9:660–664.
Article11. Poruchynsky MS, Kim JH, Nogales E, et al. Tumor cells resistant to a microtubule-depolymerizing hemiasterlin analogue, HTI-286, have mutations in alpha- or beta-tubulin and increased microtubule stability. Biochemistry. 2004. 43:13944–13954.
Article12. Chen GQ, Zhu J, Shi XG, et al. In vitro studies on cellular and molecular mechanisms of arsenic trioxide (As2O3) in the treatment of acute promyelocytic leukemia: As2O3 induces NB4 cell apoptosis with downregulation of Bcl-2 expression and modulation of PML-RAR alpha/PML proteins. Blood. 1996. 88:1052–1061.
Article13. Carré M, Carles G, André N, et al. Involvement of microtubules and mitochondria in the antagonism of arsenic trioxide on paclitaxel-induced apoptosis. Biochem Pharmacol. 2002. 63:1831–1842.
Article14. Palazzo A, Ackerman B, Gundersen GG. Cell biology: Tubulin acetylation and cell motility. Nature. 2003. 421:230.15. Infante AS, Stein MS, Zhai Y, Borisy GG, Gundersen GG. Detyrosinated (Glu) microtubules are stabilized by an ATP-sensitive plus-end cap. J Cell Sci. 2000. 113:3907–3919.
Article16. Low JA, Wedam SB, Lee JJ, et al. Phase II clinical trial of ixabepilone (BMS-247550), an epothilone B analog, in metastatic and locally advanced breast cancer. J Clin Oncol. 2005. 23:2726–2734.
Article17. Gottesman MM, Fojo T, Bates SE. Multidrug resistance in cancer: role of ATP-dependent transporters. Nat Rev Cancer. 2002. 2:48–58.
Article18. Orr GA, Verdier-Pinard P, McDaid H, Horwitz SB. Mechanisms of Taxol resistance related to microtubules. Oncogene. 2003. 22:7280–7295.
Article19. Hari M, Wang Y, Veeraraghavan S, Cabral F. Mutations in alpha- and beta-tubulin that stabilize microtubules and confer resistance to colcemid and vinblastine. Mol Cancer Ther. 2003. 2:597–605.20. Freitas RA, Silva dos, Gimenes Teixeira HL, et al. Apoptosis induction by (+)alpha-tocopheryl succinate in the absence or presence of all-trans retinoic acid and arsenic trioxide in NB4, NB4-R2 and primary APL cells. Leuk Res. 2009. 33:958–963.
Article21. Dong JT, Luo XM. Arsenic-induced DNA-strand breaks associated with DNA-protein crosslinks in human fetal lung fibroblasts. Mutat Res. 1993. 302:97–102.
Article22. Van Wijk R, Welters M, Souren JE, Ovelgonne H, Wiegant FA. Serum-stimulated cell cycle progression and stress protein synthesis in C3H10T1/2 fibroblasts treated with sodium arsenite. J Cell Physiol. 1993. 155:265–272.
Article23. Yih LH, Lee TC. Arsenite induces p53 accumulation through an ATM-dependent pathway in human fibroblasts. Cancer Res. 2000. 60:6346–6352.24. Minotti AM, Barlow SB, Cabral F. Resistance to antimitotic drugs in Chinese hamster ovary cells correlates with changes in the level of polymerized tubulin. J Biol Chem. 1991. 266:3987–3994.
Article25. MacRae TH. Tubulin post-translational modifications-enzymes and their mechanisms of action. Eur J Biochem. 1997. 244:265–278.
Article26. Jiang JD, Wang Y, Roboz J, Strauchen J, Holland JF, Bekesi JG. Inhibition of microtubule assembly in tumor cells by 3-bromoacetylamino benzoylurea, a new cancericidal compound. Cancer Res. 1998. 58:2126–2133.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- A Case of Relapsed Acute Promyleocytic Leukemia Induced Remission with Arsenic Trioxide(As2O3)
- Salvage treatments with all-trans retinoic acid and arsenic trioxide-based regimens in acute promyelocytic leukemia
- Remission induction using arsenic trioxide in acute promyelocytic leukemia
- Antitumor Effects of Arsenic Trioxide on Neuroblastoma
- Inducing Apoptosis of NCI-H157 Human Lung Carcinoma Cells via Activation of Caspase Cascade by Combination Treatment with Arsenic Trioxide and Sulindac
