Galectin-9 is Involved in Immunosuppression Mediated by Human Bone Marrow-derived Clonal Mesenchymal Stem Cells
- Affiliations
-
- 1Drug Development Program, Department of Medicine, Inha University School of Medicine, Incheon 22332, Korea.
- 2Translational Research Center, Inha University School of Medicine, Incheon 22332, Korea. sunuksong@inha.ac.kr, tgwise@inha.ac.kr
- 3Inha Research Institute for Medical Sciences of Biomedical Sciences, Inha University School of Medicine, Incheon 22332, Korea.
- 4SCM Lifescience Co. Ltd., Incheon 22332, Korea.
- KMID: 2150853
- DOI: http://doi.org/10.4110/in.2015.15.5.241
Abstract
- Bone marrow-derived mesenchymal stem cells (MSCs) have immunomodulatory properties and can suppress exaggerated pro-inflammatory immune responses. Although the exact mechanisms remain unclear, a variety of soluble factors are known to contribute to MSC-mediated immunosuppression. However, functional redundancy in the immunosuppressive properties of MSCs indicates that other uncharacterized factors could be involved. Galectin-9, a member of the beta-galactoside binding galectin family, has emerged as an important regulator of innate and adaptive immunity. We examined whether galectin-9 contributes to MSC-mediated immunosuppression. Galectin-9 was strongly induced and secreted from human MSCs upon stimulation with pro-inflammatory cytokines. An in vitro immunosuppression assay using a knockdown approach revealed that galectin-9-deficient MSCs do not exert immunosuppressive activity. We also provided evidence that galectin-9 may contribute to MSC-mediated immunosuppression by binding to its receptor, TIM-3, expressed on activated lymphocytes, leading to apoptotic cell death of activated lymphocytes. Taken together, our findings demonstrate that galectin-9 is involved in MSC-mediated immunosuppression and represents a potential therapeutic factor for the treatment of inflammatory diseases.
MeSH Terms
Figure
-
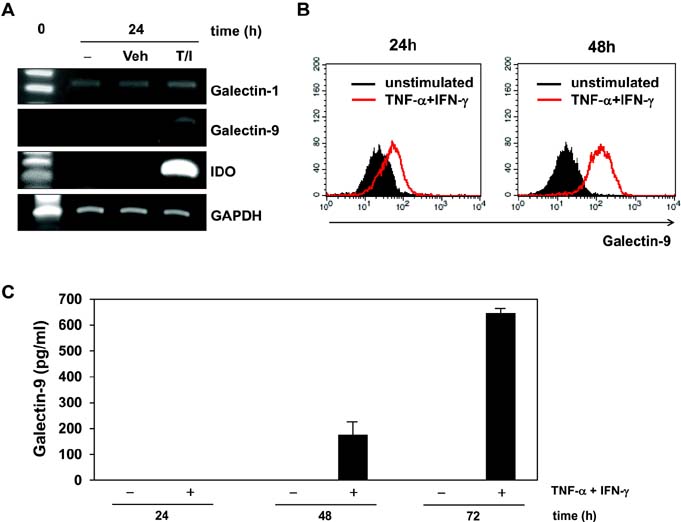
Figure 1 Galectin-9 induction in MSCs by experimental inflammatory stimulation. (A) MSCs were stimulated with 10 ng/ml TNF-α and 20 ng/ml IFN-γ for 24 h and expression of IDO, galectin-1, galectin-9 and GAPDH was detected by RT-PCR. (B) After MSCs were stimulated with 10 ng/ml TNF-α and 20 ng/ml IFN-γ for 24 and 48 h, flow cytometric analysis was conducted to detect intracellular expression of galectin-9. Red lines histograms indicate intracellular galectin-9 after stimulation while black-filled histograms represent unstimulated controls. (C) MSCs were stimulated with 10 ng/ml TNF-α and 20 ng/ml IFN-γ up to 72 h and then secreted galectin-9 in the culture supernatant was quantified by ELISA. Data represent the average of three independent experiments.
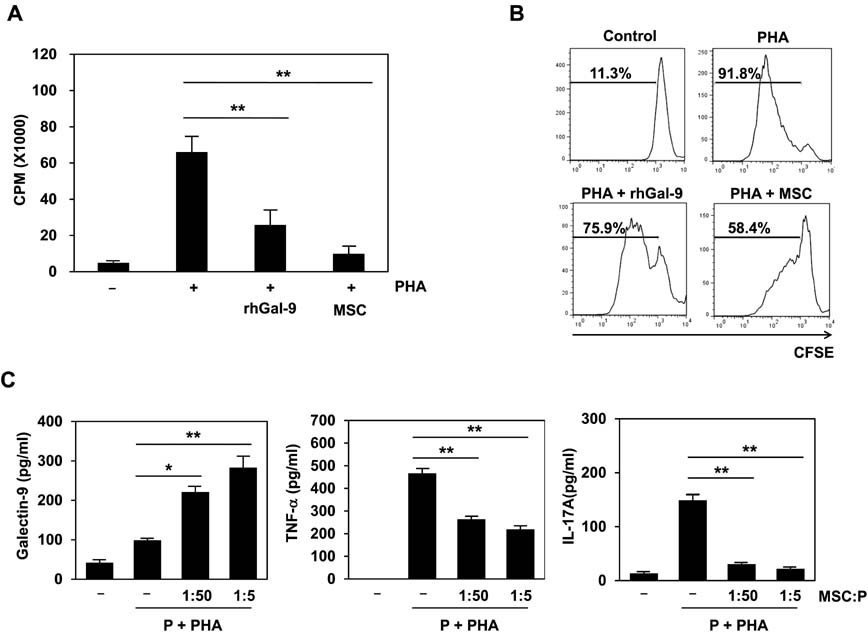
Figure 2 The immunosuppression of galectin-9 or MSCs on human lymphocyte proliferation. (A) In vitro immunosuppressive effects of galectin-9 or MSCs on human lymphocyte proliferation were evaluated by using mitogenically activated human PBMCs (P). PBMCs stimulated with PHA (1 µg/ml) were treated with 1.5 µg/ml of recombinant human galectin-9 (rhGal-9) or co-cultured with MSCs (the cell number ratio of MSC to PBMC is 1:5) for 3 days. Lymphocyte proliferation was determined by [3H]-thymidine incorporation. (B) CFSE-labeled PBMCs stimulated with PHA were incubated with 1.5 µg/ml rhGal-9 or co-cultured with MSCs (MSC:PBMC=1:5) for 5 days followed by flow cytometric analysis for cell division. Dividing PBMCs were shown in percentage. Data shown are representative of three independent experiments. (C) Galectin-9, TNF-α, and IL-17A in the culture supernatants were quantitated by ELISA. Data are the average of three independent experiments. Statistical significance was *p<0.05 or **p<0.005. P: PBMC.
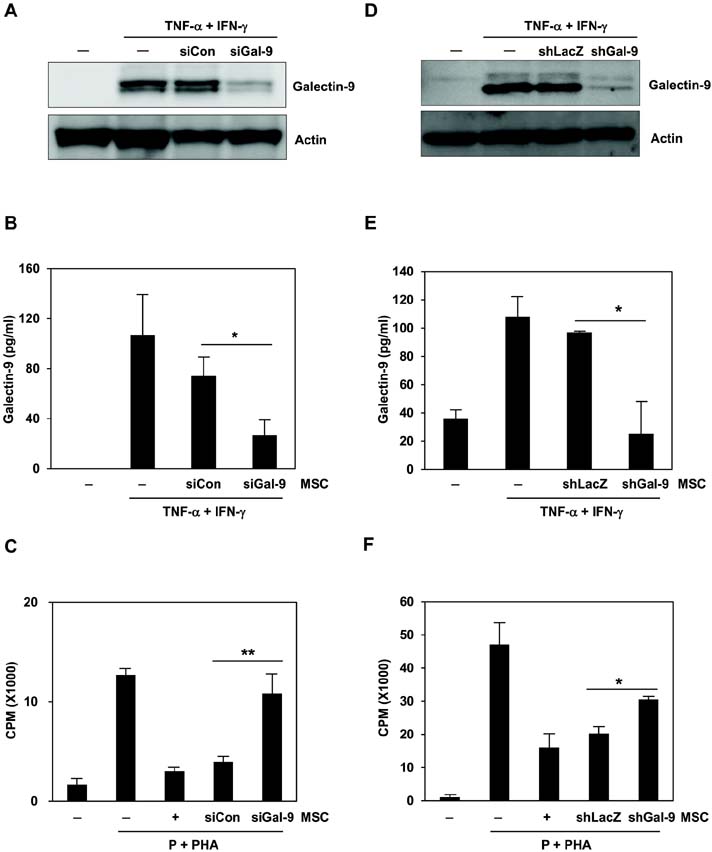
Figure 3 The effects of galectin-9-knockdown MSCs on proliferation of activated lymphocytes. (A) MSCs were transfected with siRNA oligomers specific to galectin-9 (siGal-9) or scrambled control oligomers (siCon). After the cells were stimulated with 10 ng/ml TNF-α and 20 ng/ml IFN-γ for 48 h, western blot analysis was performed to check galectin-9 knockdown. (B) siGal-9-transfected MSCs were stimulated with10 ng/ml TNF-α and 20 ng/ml IFN-γ for 48 h and then secretion of galectin-9 was determined by ELISA. (C) PBMCs stimulated with 1 µg/ml PHA were co-cultured with siCon- or siGal-9-transfected MSCs (MSC:PBMC=1:5) for 3 days. Lymphocyte proliferation was determined by [3H]-thymidine incorporation. Galectin-9 knockdown by siGal-9-transfection abrogated the immunosuppressive activity of MSCs. Data shown are the average of three independent experiments. (D) MSCs were infected with shRNA-expressing lentivirus targeting galectin-9 (shGal-9) or control LacZ virus (shLacZ). After the cells were stimulated with 10 ng/ml TNF-α and 20 ng/ml IFN-γ for 48 h, western blot analysis was performed to check galectin-9 knockdown. (E) The MSCs were stimulated with 10 ng/ml TNF-α and 20 ng/ml IFN-γ for 48 h and then soluble galectin-9 released into the culture supernatant was determined by ELISA. (F) PBMCs stimulated with 1 µg/ml PHA were co-cultured with shLacZ- or shGal-9-infected MSCs for 3 days. Lymphocyte proliferation was determined as in (C). MSC-suppressed proliferation of activated PBMCs was significantly recovered in the co-culture of shGal-9-transfected MSCs. Data are the average of three independent experiments. Statistical significance was *p< 0.05 or **p<0.005. P: PBMC.
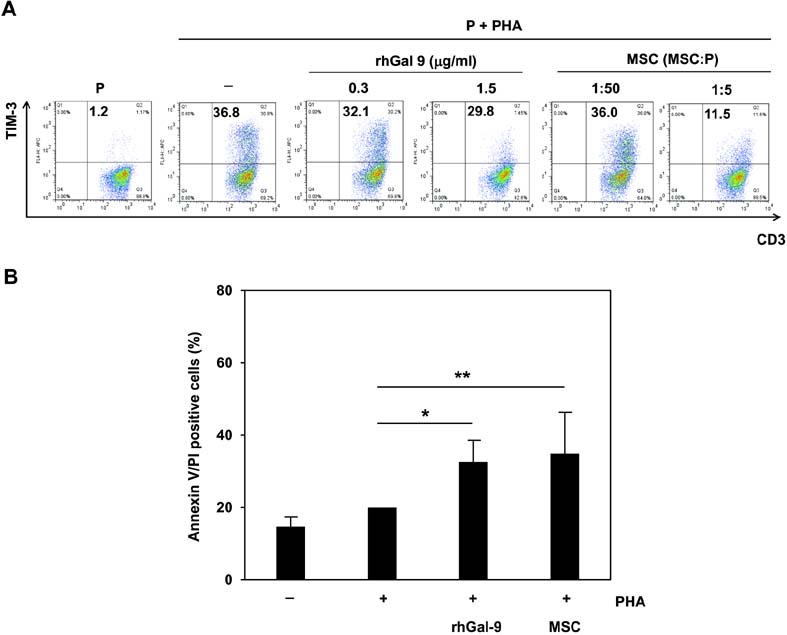
Figure 4 The effects of galectin-9 or MSCs on TIM-3-expressing lymphocytes and apoptotic analysis of activated lymphocytes by galectin-9-or MSCs. (A) After PHA-activated PBMCs were treated with exogenous rhGal-9 at a concentration of 0.3 or 1.5 µg/ml or were co-cultured with MSCs at the indicated cell ratios for 3 days, flow cytometric analysis was conducted to examine the effects of galectin-9 or MSCs on TIM-3-expressing CD3+ lymphocytes. (B) The apoptosis-inducing effects of galectin-9 or MSCs on PHA-activated PBMCs were evaluated. In the presence of rhGal-9 or MSCs, activation-induced cell death of PBMCs was significantly increased. Apoptotic cell death induced by rhGal-9 was comparable to that induced by MSCs. Apoptotic cell death was evaluated by annexin V and PI staining. Data are the average of three independent experiments. Statistical significance was *p<0.05 or **p<0.005. P: PBMC.
Reference
-
1. da Silva Meirelles L, Chagastelles PC, Nardi NB. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J Cell Sci. 2006; 119:2204–2213.
Article2. Zuk PA, Zhu M, Ashjian P, De Ugarte DA, Huang JI, Mizuno H, Alfonso ZC, Fraser JK, Benhaim P, Hedrick MH. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell. 2002; 13:4279–4295.
Article3. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science. 1999; 284:143–147.
Article4. Uccelli A, Moretta L, Pistoia V. Mesenchymal stem cells in health and disease. Nat Rev Immunol. 2008; 8:726–736.
Article5. Le Blanc K, Frassoni F, Ball L, Locatelli F, Roelofs H, Lewis I, Lanino E, Sundberg B, Bernardo ME, Remberger M, Dini G, Egeler RM, Bacigalupo A, Fibbe W, Ringden O. Mesenchymal stem cells for treatment of steroid-resistant, severe, acute graft-versus-host disease: a phase II study. Lancet. 2008; 371:1579–1586.
Article6. Jung KH, Song SU, Yi T, Jeon MS, Hong SW, Zheng HM, Lee HS, Choi MJ, Lee DH, Hong SS. Human bone marrow-derived clonal mesenchymal stem cells inhibit inflammation and reduce acute pancreatitis in rats. Gastroenterology. 2011; 140:998–1008.
Article7. Yeung TY, Seeberger KL, Kin T, Adesida A, Jomha N, Shapiro AM, Korbutt GS. Human mesenchymal stem cells protect human islets from pro-inflammatory cytokines. PLoS One. 2012; 7:e38189.
Article8. Bernardo ME, Fibbe WE. Safety and efficacy of mesenchymal stromal cell therapy in autoimmune disorders. Ann NY Acad Sci. 2012; 1266:107–117.
Article9. Stagg J, Galipeau J. Mechanisms of immune modulation by mesenchymal stromal cells and clinical translation. Curr Mol Med. 2013; 13:856–867.
Article10. Salem HK, Thiemermann C. Mesenchymal stromal cells: current understanding and clinical status. Stem Cells. 2010; 28:585–596.
Article11. English K, Ryan JM, Tobin L, Murphy MJ, Barry FP, Mahon BP. Cell contact, prostaglandin E(2) and transforming growth factor beta 1 play non-redundant roles in human mesenchymal stem cell induction of CD4+CD25 (High) forkhead box P3+ regulatory T cells. Clin Exp Immunol. 2009; 156:149–160.
Article12. Ankrum J, Karp JM. Mesenchymal stem cell therapy: Two steps forward, one step back. Trends Mol Med. 2010; 16:203–209.
Article13. Yanez R, Oviedo A, Aldea M, Bueren JA, Lamana ML. Prostaglandin E2 plays a key role in the immunosuppressive properties of adipose and bone marrow tissue-derived mesenchymal stromal cells. Exp Cell Res. 2010; 316:3109–3123.
Article14. Ryan JM, Barry F, Murphy JM, Mahon BP. Interferon-gamma does not break, but promotes the immunosuppressive capacity of adult human mesenchymal stem cells. Clin Exp Immunol. 2007; 149:353–363.
Article15. Camby I, Le MM, Lefranc F, Kiss R. Galectin-1: a small protein with major functions. Glycobiology. 2006; 16:137R–157R.
Article16. van Kooyk Y, Rabinovich GA. Protein-glycan interactions in the control of innate and adaptive immune responses. Nat Immunol. 2008; 9:593–601.
Article17. Rabinovich GA, Toscano MA. Turning 'sweet' on immunity: galectin-glycan interactions in immune tolerance and inflammation. Nat Rev Immunol. 2009; 9:338–352.
Article18. Matsushita N, Nishi N, Seki M, Matsumoto R, Kuwabara I, Liu FT, Hata Y, Nakamura T, Hirashima M. Requirement of divalent galactoside-binding activity of ecalectin/galectin-9 for eosinophil chemoattraction. J Biol Chem. 2000; 275:8355–8360.
Article19. Matsumoto R, Hirashima M, Kita H, Gleich GJ. Biological activities of ecalectin: a novel eosinophil-activating factor. J Immunol. 2002; 168:1961–1967.
Article20. Arikawa T, Saita N, Oomizu S, Ueno M, Matsukawa A, Katoh S, Kojim K, Nagahara K, Miyake M, Yamauchi A, Kohrogi H, Hirashima M. Galectin-9 expands immunosuppressive macrophages to ameliorate T-cell-mediated lung inflammation. Eur J Immunol. 2010; 40:548–558.
Article21. Irie A, Yamauchi A, Kontani K, Kihara M, Liu D, Shirato Y, Seki M, Nishi N, Nakamura T, Yokomise H, Hirashima M. Galectin-9 as a prognostic factor with antimetastatic potential in breast cancer. Clin Cancer Res. 2005; 11:2962–2968.
Article22. Kageshita T, Kashio Y, Yamauchi A, Seki M, Abedin MJ, Nishi N, Shoji H, Nakamura T, Ono T, Hirashima M. Possible role of galectin-9 in cell aggregation and apoptosis of human melanoma cell lines and its clinical significance. Int J Cancer. 2002; 99:809–816.
Article23. Kashio Y, Nakamura K, Abedin MJ, Seki M, Nishi N, Yoshida N, Nakamura T, Hirashima M. Galectin-9 induces apoptosis through the calcium-calpain-caspase-1 pathway. J Immunol. 2003; 170:3631–3636.
Article24. Zhu C, Anderson AC, Schubart A, Xiong H, Imitola J, Khoury SJ, Zheng XX, Strom TB, Kuchroo VK. The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity. Nat Immunol. 2005; 6:1245–1252.
Article25. Seki M, Oomizu S, Sakata KM, Sakata A, Arikawa T, Watanabe K, Ito K, Takeshita K, Niki T, Saita N, Nishi N, Yamauchi A, Katoh S, Matsukawa A, Kuchroo V, Hirashima M. Galectin-9 suppresses the generation of Th17, promotes the induction of regulatory T cells, and regulates experimental autoimmune arthritis. Clin Immunol. 2008; 127:78–88.
Article26. Oomizu S, Arikawa T, Niki T, Kadowaki T, Ueno M, Nishi N, Yamauchi A, Hattori T, Masaki T, Hirashima M. Cell surface galectin-9 expressing Th cells regulate Th17 and Foxp3+ Treg development by galectin-9 secretion. PLoS One. 2012; 7:e48574.
Article27. Sakai K, Kawata E, Ashihara E, Nakagawa Y, Yamauchi A, Yao H, Nagao R, Tanaka R, Yokota A, Takeuchi M, Hirai H, Kimura S, Hirashima M, Yoshimura N, Maekawa T. Galectin-9 ameliorates acute GVH disease through the induction of T-cell apoptosis. Eur J Immunol. 2011; 41:67–75.
Article28. Gieseke F, Bohringer J, Bussolari R, Dominici M, Handgretinger R, Muller I. Human multipotent mesenchymal stromal cells use galectin-1 to inhibit immune effector cells. Blood. 2010; 116:3770–3779.
Article29. Sioud M, Mobergslien A, Boudabous A, Floisand Y. Mesenchymal stem cell-mediated T cell suppression occurs through secreted galectins. Int J Oncol. 2011; 38:385–390.
Article30. Song SU, Kim CS, Yoon SP, Kim SK, Lee MH, Kang JS, Choi GS, Moon SH, Choi MS, Cho YK, Son BK. Variations of clonal marrow stem cell lines established from human bone marrow in surface epitopes, differentiation potential, gene expression, and cytokine secretion. Stem Cells Dev. 2008; 17:451–461.
Article31. Keating A. Mesenchymal stromal cells. Curr Opin Hematol. 2006; 13:419–425.
Article32. Yang RY, Rabinovich GA, Liu FT. Galectins: structure, function and therapeutic potential. Expert Rev Mol Med. 2008; 10:e17.
Article33. Di Nicola M, Carlo-Stella C, Magni M, Milanesi M, Longoni PD, Matteucci P, Grisanti S, Gianni AM. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood. 2002; 99:3838–3843.
Article34. Tse WT, Pendleton JD, Beyer WM, Egalka MC, Guinan EC. Suppression of allogeneic T-cell proliferation by human marrow stromal cells: implications in transplantation. Transplantation. 2003; 75:389–397.
Article35. Singer NG, Caplan AI. Mesenchymal stem cells: mechanisms of inflammation. Annu Rev Pathol. 2011; 6:457–478.
Article36. Yi T, Song SU. Immunomodulatory properties of mesenchymal stem cells and their therapeutic applications. Arch Pharm Res. 2012; 35:213–221.
Article37. Dorronsoro A, Fernandez-Rueda J, Fechter K, Ferrin I, Salcedo JM, Jakobsson E, Trigueros C. Human mesenchymal stromal cell-mediated immunoregulation: mechanisms of action and clinical applications. Bone Marrow Res. 2013; 2013:203643.
Article38. Sioud M. New insights into mesenchymal stromal cell-mediated T-cell suppression through galectins. Scand J Immunol. 2011; 73:79–84.
Article39. Almkvist J, Karlsson A. Galectins as inflammatory mediators. Glycoconj J. 2004; 19:575–581.
Article40. Rabinovich GA, Ramhorst RE, Rubinstein N, Corigliano A, Daroqui MC, Kier-Joffe EB, Fainboim L. Induction of allogenic T-cell hyporesponsiveness by galectin-1-mediated apoptotic and non-apoptotic mechanisms. Cell Death Differ. 2002; 9:661–670.
Article41. He J, Baum LG. Presentation of galectin-1 by extracellular matrix triggers T cell death. J Biol Chem. 2004; 279:4705–4712.
Article42. van der Leij J, van den Berg A, Blokzijl T, Harms G, van Goor H, Zwiers P, van Weeghel R, Poppema S, Visser L. Dimeric galectin-1 induces IL-10 production in T-lymphocytes: an important tool in the regulation of the immune response. J Pathol. 2004; 204:511–518.
Article43. Rabinovich GA, Ariel A, Hershkoviz R, Hirabayashi J, Kasai KI, Lider O. Specific inhibition of T-cell adhesion to extracellular matrix and proinflammatory cytokine secretion by human recombinant galectin-1. Immunology. 1999; 97:100–106.
Article44. Yang RY, Hsu DK, Liu FT. Expression of galectin-3 modulates T-cell growth and apoptosis. Proc Natl Acad Sci USA. 1996; 93:6737–6742.
Article45. Lin HM, Pestell RG, Raz A, Kim HR. Galectin-3 enhances cyclin D(1) promoter activity through SP1 and a cAMP-responsive element in human breast epithelial cells. Oncogene. 2002; 21:8001–8010.
Article46. Stillman BN, Hsu DK, Pang M, Brewer CF, Johnson P, Liu FT, Baum LG. Galectin-3 and galectin-1 bind distinct cell surface glycoprotein receptors to induce T cell death. J Immunol. 2006; 176:778–789.
Article47. Dhirapong A, Lleo A, Leung P, Gershwin ME, Liu FT. The immunological potential of galectin-1 and -3. Autoimmun Rev. 2009; 8:360–363.
Article48. Frigeri LG, Zuberi RI, Liu FT. Epsilon BP, a beta-galactoside-binding animal lectin, recognizes IgE receptor (Fc epsilon RI) and activates mast cells. Biochemistry. 1993; 32:7644–7649.
Article49. Chen HY, Sharma BB, Yu L, Zuberi R, Weng IC, Kawakami Y, Kawakami T, Hsu DK, Liu FT. Role of galectin-3 in mast cell functions: galectin-3-deficient mast cells exhibit impaired mediator release and defective JNK expression. J Immunol. 2006; 177:4991–4997.
Article50. Hastings WD, Anderson DE, Kassam N, Koguchi K, Greenfield EA, Kent SC, Zheng XX, Strom TB, Hafler DA, Kuchroo VK. TIM-3 is expressed on activated human CD4+ T cells and regulates Th1 and Th17 cytokines. Eur J Immunol. 2009; 39:2492–2501.
Article51. He W, Fang Z, Wang F, Wu K, Xu Y, Zhou H, Du D, Gao Y, Zhang WN, Niki T, Hirashima M, Yuan J, Chen ZK. Galectin-9 significantly prolongs the survival of fully mismatched cardiac allografts in mice. Transplantation. 2009; 88:782–790.
Article52. Koguchi K, Anderson DE, Yang L, O'Connor KC, Kuchroo VK, Hafler DA. Dysregulated T cell expression of TIM3 in multiple sclerosis. J Exp Med. 2006; 203:1413–1418.
Article53. Chou FC, Shieh SJ, Sytwu HK. Attenuation of Th1 response through galectin-9 and T-cell Ig mucin 3 interaction inhibits autoimmune diabetes in NOD mice. Eur J Immunol. 2009; 39:2403–2411.
Article54. Krampera M. Mesenchymal stromal cell 'licensing': a multistep process. Leukemia. 2011; 25:1408–1414.
Article55. Monney L, Sabatos CA, Gaglia JL, Ryu A, Waldner H, Chernova T, Manning S, Greenfield EA, Coyle AJ, Sobel RA, Freeman GJ, Kuchroo VK. Th1-specific cell surface protein Tim-3 regulates macrophage activation and severity of an autoimmune disease. Nature. 2002; 415:536–541.
Article56. Nakae S, Iwakura Y, Suto H, Galli SJ. Phenotypic differences between Th1 and Th17 cells and negative regulation of Th1 cell differentiation by IL-17. J Leukoc Biol. 2007; 81:1258–1268.
Article57. Gleason MK, Lenvik TR, McCullar V, Felices M, O'Brien MS, Cooley SA, Verneris MR, Cichocki F, Holman CJ, Panoskaltsis-Mortari A, Niki T, Hirashima M, Blazar BR, Miller JS. Tim-3 is an inducible human natural killer cell receptor that enhances interferon gamma production in response to galectin-9. Blood. 2012; 119:3064–3072.
Article58. Sabatos CA, Chakravarti S, Cha E, Schubart A, Sanchez-Fueyo A, Zheng XX, Coyle AJ, Strom TB, Freeman GJ, Kuchroo VK. Interaction of Tim-3 and Tim-3 ligand regulates T helper type 1 responses and induction of peripheral tolerance. Nat Immunol. 2003; 4:1102–1110.
Article59. Plumas J, Chaperot L, Richard MJ, Molens JP, Bensa JC, Favrot MC. Mesenchymal stem cells induce apoptosis of activated T cells. Leukemia. 2005; 19:1597–1604.
Article60. Zhang Y, Ma CJ, Wang JM, Ji XJ, Wu XY, Moorman JP, Yao ZQ. Tim-3 regulates pro- and anti-inflammatory cytokine expression in human CD14+ monocytes. J Leukoc Biol. 2012; 91:189–196.
Article61. Kanzaki M, Wada J, Sugiyama K, Nakatsuka A, Teshigawara S, Murakami K, Inoue K, Terami T, Katayama A, Eguchi J, Akiba H, Yagita H, Makino H. Galectin-9 and T cell immunoglobulin mucin-3 pathway is a therapeutic target for type 1 diabetes. Endocrinology. 2012; 153:612–620.
Article62. Wiersma VR, de Bruyn M, Helfrich W, Bremer E. Therapeutic potential of Galectin-9 in human disease. Med Res Rev. 2013; 33:Suppl 1. E102–E126.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Clinical Safety and Efficacy of Autologous Bone Marrow-Derived Mesenchymal Stem Cell Transplantation in Sensorineural Hearing Loss Patients
- Concise Review: Differentiation of Human Adult Stem Cells Into Hepatocyte-like Cells In vitro
- Bone Marrow-Derived Mesenchymal Stem Cells for Regenerative Medicine
- Bone marrow-derived stem cells contribute to regeneration of the endometrium
- Differential Potential of Stem Cells Following Their Origin: Subacromial Bursa, Bone Marrow, Umbilical Cord Blood
