A review on gastric leptin: the exocrine secretion of a gastric hormone
- Affiliations
-
- 1Department of Pathology and Cell Biology, University of Montreal, Montreal, Quebec, Canada. moise.bendayan@umontreal.ca
- KMID: 1447448
- DOI: http://doi.org/10.5115/acb.2012.45.1.1
Abstract
- A major advance in the understanding of the regulation of food intake has been the discovery of the adipokine leptin a hormone secreted by the adipose tissue. After crossing the blood-brain barrier, leptin reaches its main site of action at the level of the hypothalamic cells where it plays fundamental roles in the control of appetite and in the regulation of energy expenditure. At first considered as a hormone specific to the white adipose tissue, it was rapidly found to be expressed by other tissues. Among these, the gastric mucosa has been demonstrated to secrete large amounts of leptin. Secretion of leptin by the gastric chief cells was found to be an exocrine secretion. Leptin is secreted towards the gastric lumen into the gastric juice. We found that while secretion of leptin by the white adipose tissue is constitutive, secretion by the gastric cells is a regulated one responding very rapidly to secretory stimuli such as food intake. Exocrine-secreted leptin survives the hydrolytic conditions of the gastric juice by forming a complex with its soluble receptor. This soluble receptor is synthesized by the gastric cells and the leptin-leptin receptor complex gets formed at the level of the gastric chief cell secretory granules before being released into the gastric lumen. The leptin-leptin receptor upon resisting the hydrolytic conditions of the gastric juice is channelled, to the duodenum. Transmembrane leptin receptors expressed at the luminal membrane of the duodenal enterocytes interact with the luminal leptin. Leptin is actively transcytosed by the duodenal enterocytes. From the apical membrane it is transferred to the Golgi apparatus where it binds again its soluble receptor. The newly formed leptin-leptin receptor complex is then secreted baso-laterally into the intestinal mucosa to reach the blood capillaries and circulation thus reaching the hypothalamus where its action regulates food intake. Exocrine-secreted gastric leptin participates in the short term regulation of food intake independently from that secreted by the adipose tissue. Adipose tissue leptin on the other hand, regulates in the long term energy storage. Both tissues work in tandem to ensure management of food intake and energy expenditure.
MeSH Terms
-
Adipokines
Adipose Tissue
Adipose Tissue, White
Appetite
Blood-Brain Barrier
Capillaries
Chief Cells, Gastric
Dietary Sucrose
Duodenum
Eating
Energy Metabolism
Enterocytes
Gastric Juice
Gastric Mucosa
Golgi Apparatus
Hand
Hypothalamus
Intestinal Mucosa
Leptin
Membranes
Phenobarbital
Receptors, Leptin
Secretory Vesicles
Adipokines
Dietary Sucrose
Leptin
Phenobarbital
Receptors, Leptin
Figure
-
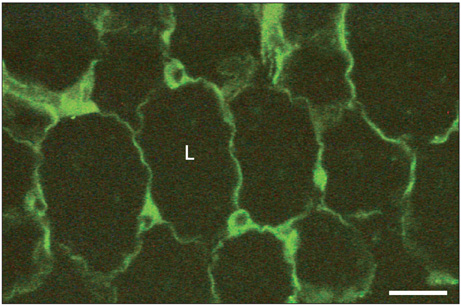
Fig. 1 Immunohistochemical detection of leptin in adipose tissue. Adipose tissue from the epididymal fat pad was fixed in Bouin's fluid and embedded in paraffin. Five µm thick tissue sections were cut and submitted to the immunohistochemical detection of leptin using an anti-leptin antibody and a fluorescein isothiocyanate (FITC)-conjugated secondary antibody. The thin cytoplasmic rim surrounding the lipid droplet (L) and the perinuclear cytoplasmic region display a specific positive green fluorescent signal revealing the presence of leptin in the adipocytes. Scale bar=50 µm.

Fig. 2 Immunocytochemical detection of leptin in adipose tissue using the electron microscopy immunogold approach. Rat epididymal white adipose tissue was fixed in glutaraldehyde and processed for embedding at -30℃ in Lowicryl K4M. Ultrathin thin tissue sections were submitted to the protein A-gold immunocytochemical technique to reveal leptin. Sections were incubated with the anti-leptin antibody followed by the protein A-gold complex (10 nm gold particles). The labeling by the gold particles is present at the level of the rough endoplasmic reticulum (RER), the Golgi apparatus (G) and some secretory vesicles. The lipid droplet (L) as well as the nucleus (N) are devoid of any signal. Few particles are seen in the extracellular space (ES). Scale bar=0.5 µm.
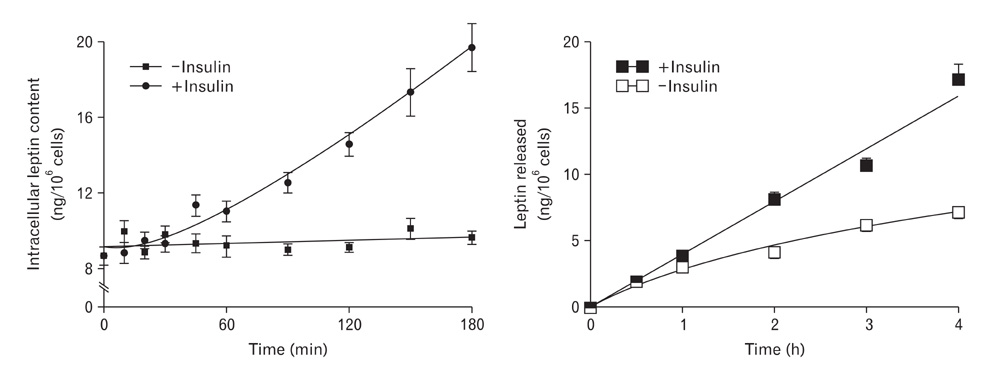
Fig. 3 Time course of leptin synthesis and secretion by isolated white adipocytes upon stimulation by insulin. Adipocytes were isolated from rat epididymal tissue by the collagenase technique and incubated in the presence or not of insulin (10 nM). Insulin is known to stimulate leptin secretion by adipocytes. Cells as well as media were sampled at regular time points and assessed for intracellular leptin content and amounts of leptin released in the culture medium. In the absence of insulin (basal condition), the amounts of leptin in the medium increase regularly over time while intracellular content of leptin remains unchanged. Upon stimulation by insulin, leptin in the medium increases much faster than under basal condition, while leptin intracellular content increases significantly reflecting stimulation of leptin synthesis by the cells. However, we can notice that upon stimulation of leptin secretion by insulin, the response of the cells is quite slow since it takes about 60 min for the increases in leptin to be significant.
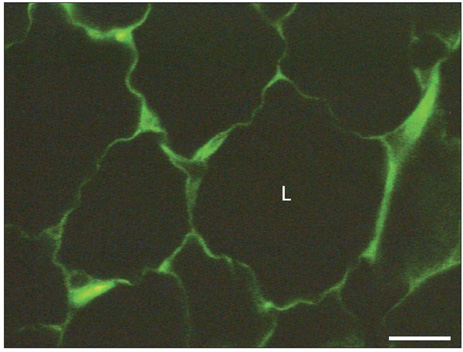
Fig. 4 Immunohistochemical detection of the leptin receptor in adipose tissue. Adipose tissue from the epididymal fat pad was fixed in Bouin's fluid and embedded in paraffin. Five µm thick tissue sections were cut and submitted to the immunohistochemical detection of the leptin receptor using an anti-leptin receptor antibody and a fluorescein isothiocyanate (FITC)-conjugated secondary antibody. Similar to the leptin signal (Fig. 1), the thin cytoplasmic rim surrounding the lipid droplet (L) and the perinuclear cytoplasmic region display a specific positive green fluorescent signal revealing the presence of the leptin receptor in the adipocytes. Scale bar=25 µm.
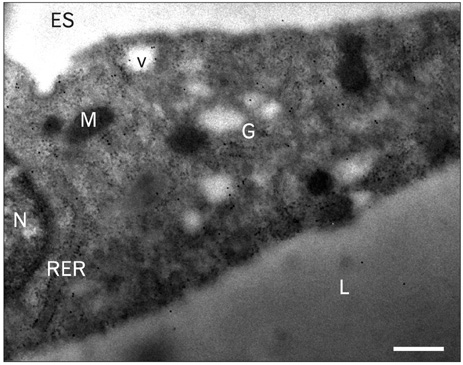
Fig. 5 Immunocytochemical detection of the leptin receptor in adipose tissue using electron microscopy and the immunogold approach. Rat epididymal white adipose tissue fixed in glutaraldehyde and processed for embedding in Lowicryl K4M. Ultrathin tissue thin sections were submitted to the protein A-gold immunocytochemical technique to reveal the leptin receptor. Sections were incubated with the anti-leptin receptor antibody followed by the protein A-gold complex (10 nm gold particles). The labeling by the gold particles is present at the level of the rough endoplasmic reticulum (RER), the Golgi apparatus (G) and some secretory vesicles (v). Very few gold particles can be detected at the level of the lipid droplet (L), the nucleus (N) or the extracellular space (ES). M, mitochondria. Scale bar=0.25 µm.
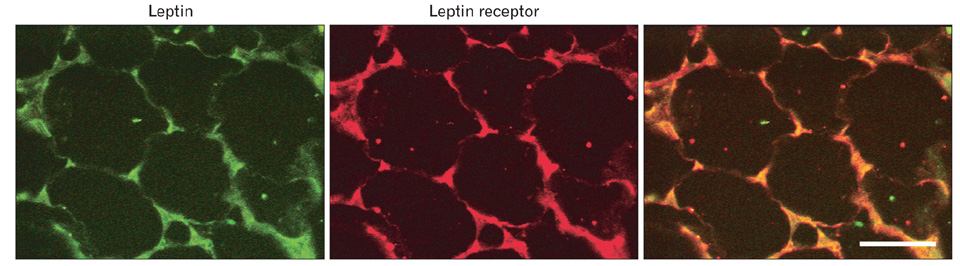
Fig. 6 Colocalization of leptin and its own receptor in the same adipocytes. Rat epididymal white adipose tissue 5 µm tissue sections was incubated with the rabbit-anti-leptin antibody and the goat-anti-leptin receptor antibody followed by an anti-rabbit IgG tagged with fluorescein isothiocyanate (FITC) and an anti-goat IgG tagged with tetramethylrhodamine isothiocyanate (TRITC) antibodies. The thin cytoplasmic rim surrounding the lipid droplet and the perinuclear regions display positive signals for both proteins; green for leptin and red for the leptin receptor. Colocalization in yellow confirms that both proteins are present in the same cellular areas. Scale bar=50 µm.

Fig. 7 (A) Gastric mucosa. Immunohistochemical detection of leptin and pepsinogen. Rat gastric mucosa was fixed in Bouin's fluid and embedded in paraffin. The 5 µm tissue sections were incubated with a rabbit anti-leptin antibody and a goat anti-pepsinogen antibody followed by an anti-rabbit IgG tagged with fluorescein isothiocyanate (FITC) and an anti-goat IgG tagged with tetramethylrhodamine isothiocyanate (TRITC) antibodies. The leptin-containing cells were revealed by the green fluorescent FITC stain while the pepsinogen-containing cells were revealed by the red fluorescent TRITC stain. We can see that the majority of the epithelial cells located in the lower half of the mucosa display positive signals for both antigens. Co-localisation confirmed the presence of both antigens in the same cells. However, close examination of the tissue sections indicates that some isolated cells (arrows) located in the lamina propria remain green, which indicates that these cells do contain leptin but are devoid of pepsinogen. This together with their location in the lamina propria suggest that these cells are pure leptin containing endocrine cells which differ from the other leptin positive cells that are epithelial chief cells from the gastric mucosa. (B) Electron microscopy of an endocrine leptin-secreting cell located in the lamina propria of the gastric mucosa of the rat. The cell is neighbouring a gastric epithelial cell and contains a large number of secretory granules. At high magnification we can identify those secretory granules as containing leptin. Indeed upon application of the immunogold approach with an anti-leptin antibody and the protein A-gold, the secretory granules display numerous gold particles revealing the presence of leptin in the granules. In a different set of experiments, using an anti-leptin receptor antibody together with the protein A-gold we were able to demonstrate that the endocrine leptin secreting cells also secrete the leptin receptor. The presence of numerous gold particles in the granules demonstrates that these granules contain both the leptin and the leptin receptor. M, mitochondria; g, secretory granules. Scale bars=50 µm (A), 1 µm (upper panel in B), 0.5 µm (bottom panel, left column), 0.25 µm (bottom panel, right column).

Fig. 8 Human gastric mucosa. Immunolocalization of leptin, pepsinogen and lipase. The semi-thin sections of the human gastric tissue fixed in glutaral dehyde and embedded in Epon were incubated sequentially with an anti-leptin and an anti-pepsi no gen antibody followed by the fluorescein isothiocyanate (FITC) and tetrame thylrhodamine isothiocyanate (TRITC) tagged secondary antibodies. In the second series of experiments serial tissue sections were incubated sequentially with the anti-leptin and the anti-lipase antibodies followed by the FITC and TRITC tagged secondary corresponding antibodies. Merging the two series of stainings reveals that cells containing leptin are the same as those containing pepsinogen and lipase. These are the chief cells lining part of the gastric epithelium. Scale bars=20 µm.

Fig. 9 Electron microscopy of the gastric chief cells. Immunocytochemistry revealing the presence of leptin in the secretory pathway of the chief cells. The thin tissue sections of the rat gastric mucosa fixed in glutaraldehyde and embedded in Epon were incubated with the anti-leptin antibody followed by the protein A-god complex (10 nm gold particles). The labeling by god particles is found at the level of the rough endoplasmic reticulum, the Golgi apparatus (G) and in the secretory granules (g ). Labeling is also found over the extracellular material of the gastric lumen (L). Scale bars=1 µm (left column), 2 µm (right column).
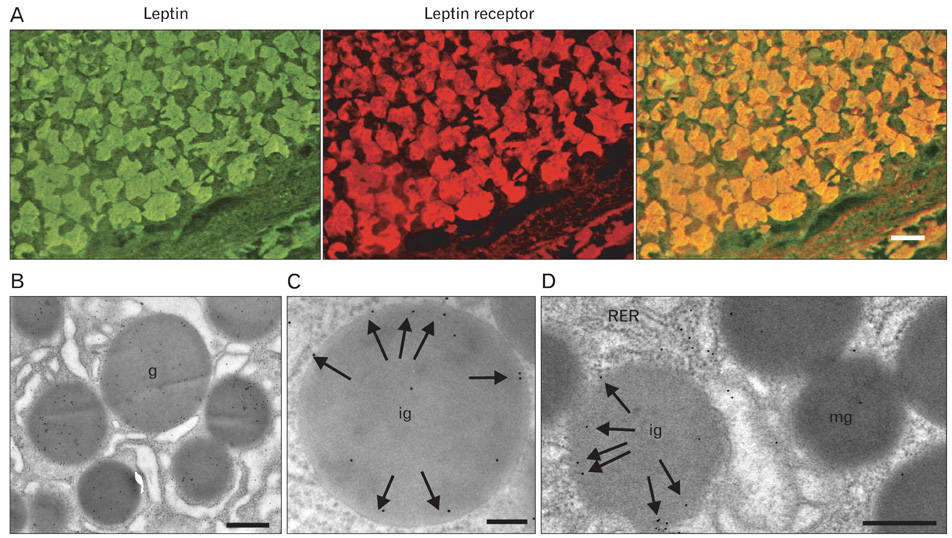
Fig. 10 (A) Leptin receptor immunohistochemistry. Rat gastric mucosa. The 5 µm tissue section was incubated sequentially with the rabbit anti-leptin antibody and the anti-leptin receptor antibody followed by the respective secondary antibodies tagged with fluorescein isothiocyanate (FITC) or tetramethylrhodamine isothiocyanate (TRICT). The stainings demonstrate the presence of leptin and the leptin receptor in the gastric mucosa chief cells. Merging the signals demonstrated that the same epithelial chief cells express both the leptin and the leptin receptor. (B) Electron microscopy immunocytochemistry. The use of the anti-leptin receptor antibody with the protein A-gold complex revealed the presence of the leptin receptor at the level of the secretory granules of the gastric chief cells. (C) When looking at the different secretory granules of a chief cell and at the distribution of the protein A-gold labeling for the leptin receptor we realize that the location of the gold particles differs between immature (ig) and mature (g) granules. While the gold particles revealing the leptin receptor are rather located towards the periphery of the immature granules, they are more centrally located in the mature ones (mg). (D) When the labeling is performed using an anti-leptin receptor monoclonal antibody directed against the trans-membrane-intracellular domain of the receptor molecule, we found that the labeling is only present in immature granules (ig) while the mature granules (mg) are rather devoid of labeling. In addition the labeling found over the immature granules is located at the periphery of the granules. This labeling disappears as the granules mature. The trans-membrane leptin receptor molecule is cleaved to generate the soluble isoform of the receptor that will bind to the leptin molecules present in the granules. RER, rough endoplasmic reticulum. Scale bars=50 µm (A), 1 µm (B), 0.5 µm (C, D).
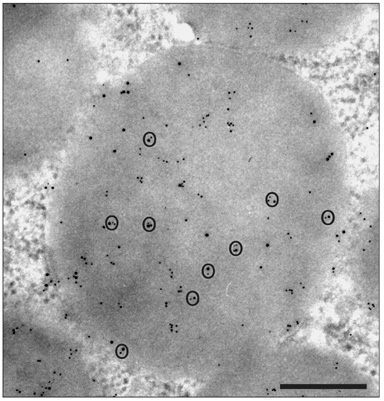
Fig. 11 Double labeling experiment. Simultaneous detection of leptin and leptin receptor in the same chief cell secretory granule. The thin tissue section was incubated with anti-leptin antibody followed by the appropriate anti-IgG molecules tagged with 5 nm gold particles. This was still followed by incubations with an anti-leptin receptor antibody followed by the appropriate anti IgG molecules tagged with 10 nm god particles. Both molecules were revealed in the same granule. In addition we can observe that small and large gold particles are often very close to each other indicating colse colocalization of leptin with its receptor and the formation of the complex. Scale bar=0.5 µm.

Fig. 12 Schematic illustration of the maturation process of the leptin receptor, the generation of the soluble isoform of the leptin receptor and the formation of the leptin-leptin receptor complex. The cell generates the long trans-membrane isoform of the leptin receptor that reaches the secretory granules within its delineating membrane. The secretory granule of the chief cells contains a large number of proteins and enzymes. We have detected pepsinogen and lipase that will be secreted to compose the gastric juice. In addition, there are convertases in the granule such as furin and proconvertase 7 (PC7). The trans-membrane long isoform of the leptin receptor is cleaved from the membrane and converted into the soluble isoform by the combined action of furin and PC7. Upon the action of both convertases, the soluble isoform is generated and released into the granule content. Once released, it binds the leptin molecules already present in the granule to form the leptin-leptin-receptor complex that will eventually be discharged by the cell into the gastric juice.
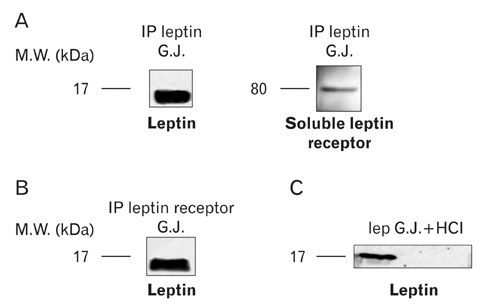
Fig. 13 Presence of the soluble leptin receptor in the gastric juice. (A) The immunoprecipitation of the rat gastric juice was carried out using an antibody against leptin. By immunoblot, we obtained one band at 16 kDa that corresponds to the leptin. We were able to reveal a second band at 80 kDa on the same immunoprecipitation, using an antibody against the extracellular portion of the leptin receptor. (B) The immunoprecipitation (IP) of the gastric juice was carried out with an antibody directed against the leptin receptor extracellular domain. By immunoblot we obtained a band at 16 kDa with the anti-leptin antibody thus confirming the association of the leptin to its soluble receptor in the gastric juice. (C) In this particular experiment, we removed the endogenous leptin-leptin receptor complex from the gastric juice by immunoprecipitation. We then added recombinant free leptin to the gastric juice and adjusted the pH to acidic conditions (pH=2), conditions mimicking those of the gastric juice in situ. An incubation at 37℃ for 30 min was carried out. The added leptin devoid of its receptor did not resist the conditions of the gastric juice and was rapidly destroyed. This demonstrates that the soluble leptin receptor is required for the leptin to form the complex in order to survive the gastric juice conditions. G.J., gastric juice.

Fig. 14 (A) Presence of the leptin receptor on the luminal membrane of the rat duodenal wall. Immunohistochemical detection was carried out using an anti leptin receptor antibody followed by a secondary antibody tagged with fluorescein isothiocyanate (FITC). The staining in the form of a thin fluorescent line is present at the apical membrane in contact with the lumen (L) of the duodenum. A much fainter and larger positive signal is also present on the basal side of the epithelial cells. (B) The presence of the leptin receptor at the apical membrane of the duodenal enterocytes has been demonstrated by electron microscopy. Applying the anti-leptin receptor antibody together with the protein A-gold on duodenal tissue sections, led to a labeling by gold particles at the level of the enterocytes apical membrane microvilli (mv) thus confirming the presence of the receptor on the luminal membrane. Scale bars=50 µm (A), 0.5 µm (B).
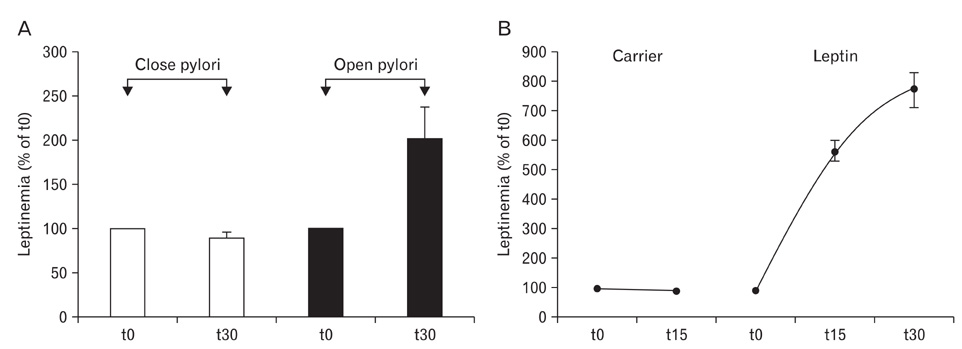
Fig. 15 Transfer of luminal gastric leptin from the duodenal lumen to the blood circulation. (A) Secretion of gastric leptin was triggered by injecting the cholinergic agonist, carbachol to anaesthetized rats. In one series of animals the pyloric sphincter was kept open while in a second series of animals, the pyloric sphincter was closed. Thirty minutes after injecting carbachol, blood was sampled and levels of leptin analyzed. When the pyloric sphincter remained open, levels of circulating leptin rise rapidly, about twice the basal level. However, when the pyloric sphincter was clamped, circulating leptin remained at basal levels. This indicates that upon stimulation, leptin secreted by the gastric cells into the gastric juice was channelled towards the duodenal lumen, absorbed by the epithelial cells and transferred to the basal side reaching the circulation along a transcytotic pathway. (B) In a second set of experiments leptin tagged with fluorescein isothiocyanate (FITC) was introduced into the lumen of the duodenum of anesthetised rats. Blood was sampled at 0, 15 and 30 min and analyzed for leptin levels by an Elisa test, demonstrating that levels of circulating leptin rise along with time. The FITC-tagged leptin was revealed by immunoblot to demonstrate that the leptin-FITC reach the circulation. This demonstrates that the rise in leptin levels are due to increasing amounts of exogenous leptin-FITC in circulation. These experiments indicate that the duodenal mucosa is able to transport leptin from the intestinal lumen to the blood stream.
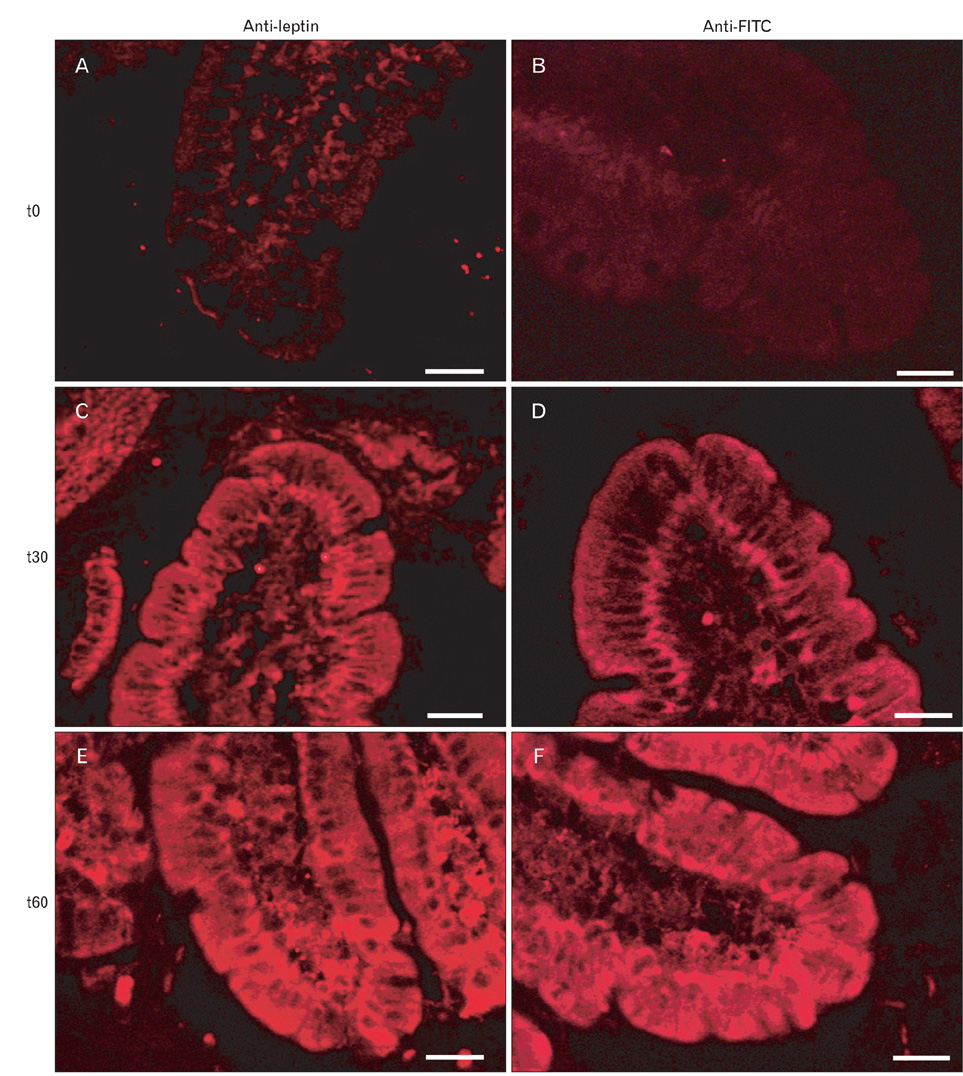
Fig. 16 Same experiment as depicted in Fig. 15B. In this case, the duodenal tissue was sampled at times 0, 30 and 60 min, fixed in Bouin's fluid and processed for immunohistochemistry. One series of tissue sections was incubated with the anti-leptin antibody followed by the secondary antibody tagged to tetramethylrhodamine isothiocyanate (TRITC). The second series of tissue sections were incubated with an anti-fluorescein isothiocyanate (FITC) antibody followed by the secondary antibody tagged to TRITC. Both series of experiments yield the same results illustrating the fact that leptin-FITC is absorbed by the duodenal epithelial cells and transported to the baso-lateral side and released into the interstitial space. At time 0, the anti-leptin antibody yielded a faint signal on the tissue while no staining was registered with the anti-FITC. At time 15 min both the anti-leptin and the anti FITC yield signals that were strong on the luminal and basal sides and weaker inside the cells. At 60 min the signals were enhanced in the cells and in the interstitial space. (A, C, E) Anti-leptin. (B, D, F) Anti-FITC. Scale bars=50 µm.
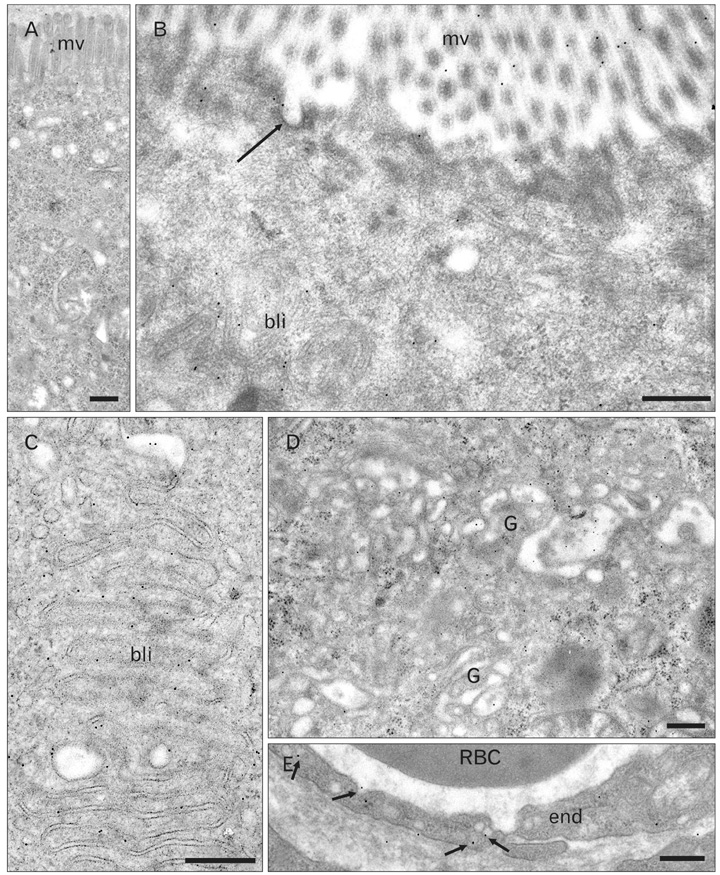
Fig. 17 Electron microscopy immunocytochemistry. Pathway of leptin-fluorescein isothiocyanate (FITC) across the duodenal epithelial cells. Same experiment as depicted in Figs. 15B and 16. The duodenal tissue was sampled 60 min after exposure to the leptin-FITC and prepared for electron microscopy. Thin tissue sections were incubated with the anti-FITC antibodies followed by the protein A-gold complex (10 nm gold particles). (A) Control condition, the duodenal tissue was not exposed to the leptin-FITC. No specific labeling is present on the tissue section. (B-E) Labeling by gold particles is found at the level of the apical membrane microvilli (mv) and in some apical membrane invaginations (arrow) corresponding to endocytotic vesicles (B). Labeling is pre sent over the baso-lateral membrane particularly at the level of the interdigitations (bli) (B, C), over the cisternae of the Golgi apparatus (G) (D) and also in the basal interstitial tissue and in the plasmalemmal vesicles of the blood capillary endothelial cells (arrows) (E). End, endothelial cells; RBC, red blood cell in capillary lumen. Scale bars=0.5 µm (A-E).
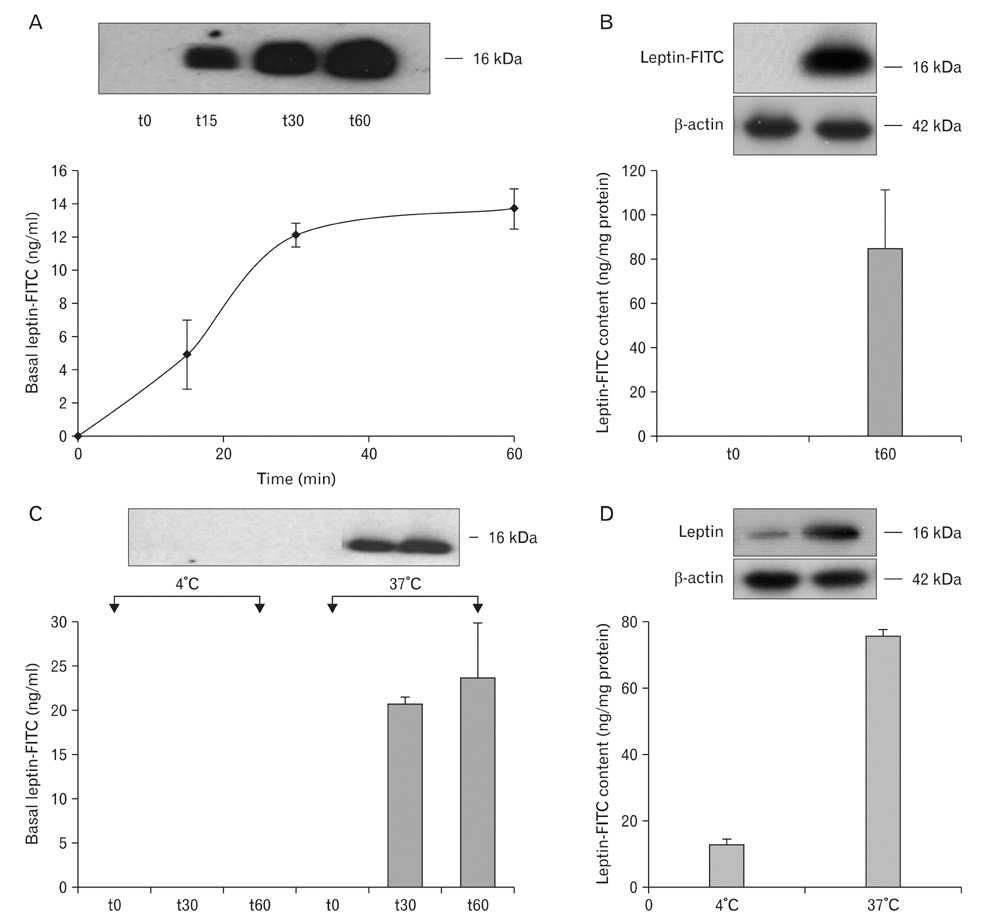
Fig. 18 Study performed in vitro. Human Caco-2 cells which represent a model for intestinal tissue were grown in vitro. These cells differentiate in a polarized epithelial monolayer. They developed tight monolayers. The cells exhibit characteristic apical microvilli and lateral junctional complexes. Cells were grown in small tight wells with separated apical and basal chambers. Leptin-fluorescein isothiocyanate (FITC) was introduced in the apical chamber and the medium of the basal chamber was sampled at different time points. Leptin-FITC appeared in the basal medium in a time-dependant and saturable manner reaching a plateau at about 40 min (A). Leptin-FITC content in the cell, absent at time 0, reached about 80 ng/mg after 60 min. (B). Carrying the experiment at 4℃ prevents the transport of the leptin-FITC across the cells (C). The uptake and internalization of the leptin-FITC by the cells were drastically reduced when the experiment was carried out at 4℃ since cell content was significantly reduced (D).

Fig. 19 Schematic drawing that illustrates the secretion of leptin by the adipocyte and the gastric chief cell. It also illustrates the fact that both types of cell secrete the leptin receptor. Both the leptin and the leptin receptor are synthesized in the rough endoplasmic reticulum, transferred to the Golgi apparatus and packaged into either small vesicles (adipocytes) or secretory granules (gastric cells). At the level of the trans-cisternae of the Golgi and in the secretory granules leptin binds its soluble receptor to form the leptin-leptin receptor complex. This complex is discharged by both cell types through an exocytotic event. The adipose tissue secretes towards the blood circulation while the gastric cells secrete in an exocrine fashion into the gastric juice. Leptin in the gastric juice is vehiculated to the duodenal lumen. In the duodenum leptin interacts with the leptin receptor present on the apical membrane of the duodenal enterocytes and is internalized by the cells. Within the intestinal epithelial cell, the leptin-leptin receptor complex is channelled within the early endosomal compartment and both molecules get separated. The leptin is channelled towards the trans-Golgi cisternae were it binds a newly synthesized soluble leptin receptor. This complex is then packaged and discharged on the baso-lateral membrane to be released into the duodenal basal interstitial space to reach the blood circulation across the capillary endothelial cells. Thus, both the gastric and the adipose tissue leptins reach the blood and circulate as a leptin-leptin receptor complex. It is under this complexed form that it reaches the target hypothalamic cells. RER, rough endoplasmic reticulum; CNS, central nervous system; N, nucleus.
Reference
-
1. Kennedy GC. The role of depot fat in the hypothalamic control of food intake in the rat. Proc R Soc Lond B Biol Sci. 1953. 140:578–596.2. Hervey GR. The effects of lesions in the hypothalamus in parabiotic rats. J Physiol. 1959. 145:336–352.3. Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994. 372:425–432.4. Tartaglia LA, Dembski M, Weng X, Deng N, Culpepper J, Devos R, Richards GJ, Campfield LA, Clark FT, Deeds J, Muir C, Sanker S, Moriarty A, Moore KJ, Smutko JS, Mays GG, Wool EA, Monroe CA, Tepper RI. Identification and expression cloning of a leptin receptor, OB-R. Cell. 1995. 83:1263–1271.5. Chen H, Charlat O, Tartaglia LA, Woolf EA, Weng X, Ellis SJ, Lakey ND, Culpepper J, Moore KJ, Breitbart RE, Duyk GM, Tepper RI, Morgenstern JP. Evidence that the diabetes gene encodes the leptin receptor: identification of a mutation in the leptin receptor gene in db/db mice. Cell. 1996. 84:491–495.6. Lee GH, Proenca R, Montez JM, Carroll KM, Darvishzadeh JG, Lee JI, Friedman JM. Abnormal splicing of the leptin receptor in diabetic mice. Nature. 1996. 379:632–635.7. Lammert A, Kiess W, Bottner A, Glasow A, Kratzsch J. Soluble leptin receptor represents the main leptin binding activity in human blood. Biochem Biophys Res Commun. 2001. 283:982–988.8. Yang G, Ge H, Boucher A, Yu X, Li C. Modulation of direct leptin signaling by soluble leptin receptor. Mol Endocrinol. 2004. 18:1354–1362.9. Liebling DS, Eisner JD, Gibbs J, Smith GP. Intestinal satiety in rats. J Comp Physiol Psychol. 1975. 89:955–965.10. Guilmeau S, Buyse M, Tsocas A, Laigneau JP, Bado A. Duodenal leptin stimulates cholecystokinin secretion: evidence of a positive leptin-cholecystokinin feedback loop. Diabetes. 2003. 52:1664–1672.11. Baggio LL, Drucker DJ. Biology of incretins: GLP-1 and GIP. Gastroenterology. 2007. 132:2131–2157.12. Ahima RS, Antwi DA. Brain regulation of appetite and satiety. Endocrinol Metab Clin North Am. 2008. 37:811–823.13. Unniappan S, Kieffer TJ. Leptin extends the anorectic effects of chronic PYY(3-36) administration in ad libitum-fed rats. Am J Physiol Regul Integr Comp Physiol. 2008. 295:R51–R58.14. Jeong KJ, Lee SY. High-level production of human leptin by fed-batch cultivation of recombinant Escherichia coli and its purification. Appl Environ Microbiol. 1999. 65:3027–3032.15. Zheng D, Jones JP, Usala SJ, Dohm GL. Differential expression of ob mRNA in rat adipose tissues in response to insulin. Biochem Biophys Res Commun. 1996. 218:434–437.16. Zheng D, Wooter MH, Zhou Q, Dohm GL. The effect of exercise on ob gene expression. Biochem Biophys Res Commun. 1996. 225:747–750.17. Cammisotto PG, Gingras D, Renaud C, Levy E, Bendayan M. Secretion of soluble leptin receptors by exocrine and endocrine cells of the gastric mucosa. Am J Physiol Gastrointest Liver Physiol. 2006. 290:G242–G249.18. Cammisotto PG, Bukowiecki LJ. Mechanisms of leptin secretion from white adipocytes. Am J Physiol Cell Physiol. 2002. 283:C244–C250.19. Cammisotto PG, Bukowiecki LJ, Deshaies Y, Bendayan M. Leptin biosynthetic pathway in white adipocytes. Biochem Cell Biol. 2006. 84:207–214.20. Cammisotto PG, Levy E, Bukowiecki LJ, Bendayan M. Crosstalk between adipose and gastric leptins for the control of food intake and energy metabolism. Prog Histochem Cytochem. 2010. 45:143–200.21. Cammisotto PG, Renaud C, Gingras D, Delvin E, Levy E, Bendayan M. Endocrine and exocrine secretion of leptin by the gastric mucosa. J Histochem Cytochem. 2005. 53:851–860.22. Bado A, Levasseur S, Attoub S, Kermorgant S, Laigneau JP, Bortoluzzi MN, Moizo L, Lehy T, Guerre-Millo M, Le Marchand-Brustel Y, Lewin MJ. The stomach is a source of leptin. Nature. 1998. 394:790–793.23. Cinti S, Matteis RD, Picó C, Ceresi E, Obrador A, Maffeis C, Oliver J, Palou A. Secretory granules of endocrine and chief cells of human stomach mucosa contain leptin. Int J Obes Relat Metab Disord. 2000. 24:789–793.24. Lindqvist A, de la Cour CD, Stegmark A, Håkanson R, Erlanson-Albertsson C. Overeating of palatable food is associated with blunted leptin and ghrelin responses. Regul Pept. 2005. 130:123–132.25. Sánchez J, Oliver P, Palou A, Picó C. The inhibition of gastric ghrelin production by food intake in rats is dependent on the type of macronutrient. Endocrinology. 2004. 145:5049–5055.26. Sobhani I, Bado A, Vissuzaine C, Buyse M, Kermorgant S, Laigneau JP, Attoub S, Lehy T, Henin D, Mignon M, Lewin MJ. Leptin secretion and leptin receptor in the human stomach. Gut. 2000. 47:178–183.27. Sobhani I, Buyse M, Goïot H, Weber N, Laigneau JP, Henin D, Soul JC, Bado A. Vagal stimulation rapidly increases leptin secretion in human stomach. Gastroenterology. 2002. 122:259–263.28. Morton NM, Emilsson V, Liu YL, Cawthorne MA. Leptin action in intestinal cells. J Biol Chem. 1998. 273:26194–26201.29. Breidert M, Miehlke S, Glasow A, Orban Z, Stolte M, Ehninger G, Bayerdörffer E, Nettesheim O, Halm U, Haidan A, Bornstein SR. Leptin and its receptor in normal human gastric mucosa and in Helicobacter pylori-associated gastritis. Scand J Gastroenterol. 1999. 34:954–961.30. Barrenetxe J, Villaro AC, Guembe L, Pascual I, Muñoz-Navas M, Barber A, Lostao MP. Distribution of the long leptin receptor isoform in brush border, basolateral membrane, and cytoplasm of enterocytes. Gut. 2002. 50:797–802.31. Cammisotto PG, Gingras D, Bendayan M. Transcytosis of gastric leptin through the rat duodenal mucosa. Am J Physiol Gastrointest Liver Physiol. 2007. 293:G773–G779.32. Schneeman BO. Gastrointestinal physiology and functions. Br J Nutr. 2002. 88:Suppl 2. S159–S163.33. Ducroc R, Guilmeau S, Akasbi K, Devaud H, Buyse M, Bado A. Luminal leptin induces rapid inhibition of active intestinal absorption of glucose mediated by sodium-glucose cotransporter 1. Diabetes. 2005. 54:348–354.34. El Homsi M, Ducroc R, Claustre J, Jourdan G, Gertler A, Estienne M, Bado A, Scoazec JY, Plaisancié P. Leptin modulates the expression of secreted and membrane-associated mucins in colonic epithelial cells by targeting PKC, PI3K, and MAPK pathways. Am J Physiol Gastrointest Liver Physiol. 2007. 293:G365–G373.35. Buyse M, Berlioz F, Guilmeau S, Tsocas A, Voisin T, Péranzi G, Merlin D, Laburthe M, Lewin MJ, Rozé C, Bado A. PepT1-mediated epithelial transport of dipeptides and cephalexin is enhanced by luminal leptin in the small intestine. J Clin Invest. 2001. 108:1483–1494.36. Stan S, Levy E, Bendayan M, Zoltowska M, Lambert M, Michaud J, Asselin C, Delvin EE. Effect of human recombinant leptin on lipid handling by fully differentiated Caco-2 cells. FEBS Lett. 2001. 508:80–84.37. Igel M, Lindenthal B, Giesa U, von BK. Evidence that leptin contributes to intestinal cholesterol absorption in obese (ob/ob) mice and wild-type mice. Lipids. 2002. 37:153–157.38. Bendayan M, Ziv E, Ben-Sasson R, Bar-On H, Kidron M. Morpho-cytochemical and biochemical evidence for insulin absorption by the rat ileal epithelium. Diabetologia. 1990. 33:197–204.39. Bruneau N, Bendayan M, Gingras D, Ghitescu L, Levy E, Lombardo D. Circulating bile salt-dependent lipase originates from the pancreas via intestinal transcytosis. Gastroenterology. 2003. 124:470–480.40. Cloutier M, Gingras D, Bendayan M. Internalization and transcytosis of pancreatic enzymes by the intestinal mucosa. J Histochem Cytochem. 2006. 54:781–794.41. Cammisotto PG, Bendayan M, Sané A, Dominguez M, Garofalo C, Levy E. Receptor-mediated transcytosis of leptin through human intestinal cells in vitro. Int J Cell Biol. 2010. 2010:928169.42. Halaas JL, Gajiwala KS, Maffei M, Cohen SL, Chait BT, Rabinowitz D, Lallone RL, Burley SK, Friedman JM. Weight-reducing effects of the plasma protein encoded by the obese gene. Science. 1995. 269:543–546.43. Pelleymounter MA, Cullen MJ, Baker MB, Hecht R, Winters D, Boone T, Collins F. Effects of the obese gene product on body weight regulation in ob/ob mice. Science. 1995. 269:540–543.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Relevance of Plasma Leptin Levels to Dysmotility-Like Functional Dyspepsia
- Immunohistochemical Expression of Neuron Specific Enolase-Positive Cells in Gastric Adenocarcinomas
- Hypercalcemia as Initial Presentation of Metastatic Adenocarcinoma of Gastric Origin: A Case Report and Review of the Literature
- Ghrelin; Influences on Helicobacter pylori-associated Gastric Diseases
- Correlation between the Serum Leptin Level and the Expression of Leptin in Stomach Cancer Patients
