A Potential Therapy Using Engineered Stem Cells Prevented Malignant Melanoma in Cellular and Xenograft Mouse Models
- Affiliations
-
- 1Laboratory of Biochemistry and Immunology, College of Veterinary Medicine, Chungbuk National University, Cheongju, Korea. kchoi@cbu.ac.kr
- 2Department of Medicine, Faculty of Medicine, University of British Columbia, Vancouver, BC, Canada.
- 3Institute of Life Science and Bio-Engineering, TheraCell Bio & Science, Cheongju, Korea.
- KMID: 2464425
- DOI: http://doi.org/10.4143/crt.2018.364
Abstract
- PURPOSE
In the present study, human neural stem cells (hNSCs) with tumor-tropic behavior were used as drug delivery vehicle to selectively target melanoma. A hNSC line (HB1.F3) was transduced into two types: one expressed only the cytosine deaminase (CD) gene (HB1.F3. CD) and the other expressed both CD and human interferon-β (IFN-β) genes (HB1.F3.CD. IFN-β).
MATERIALS AND METHODS
This study verified the tumor-tropic migratory competence of engineered hNSCs on melanoma (A375SM) using a modified Boyden chamber assay in vitro and CM-DiI staining in vivo. The antitumor effect of HB1.F3.CD and HB1.F3.CD.IFN-β on melanoma was also confirmed using an MTT assay in vitro and xenograft mouse models.
RESULTS
A secreted form of IFN-β from the HB1.F3.CD.IFN-β cells modified the epithelial-mesenchymal transition (EMT) process and metastasis of melanoma. 5-Fluorouracil treatment also accelerated the expression of the pro-apoptotic protein BAX and decelerated the expression of the anti-apoptotic protein Bcl-xL on melanoma cell line.
CONCLUSION
Our results illustrate that engineered hNSCs prevented malignant melanoma cells from proliferating in the presence of the prodrug, and the form that secreted IFN-β intervened in the EMT process and melanoma metastasis. Hence, neural stem cell-directed enzyme/prodrug therapy is a plausible treatment for malignant melanoma.
MeSH Terms
Figure
-
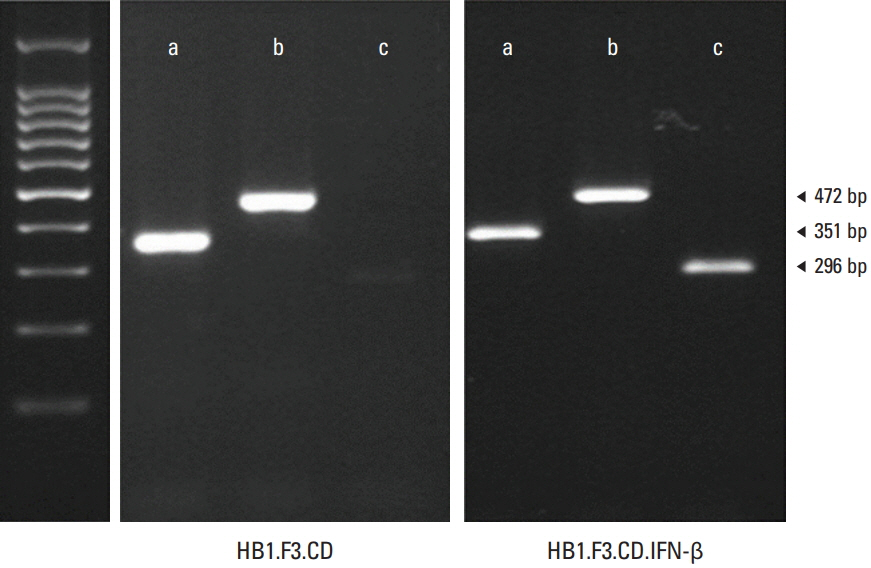
Fig. 1. Expression of Escherichia coli cytosine deaminase (CD) and human interferon β (IFN-β) gene in engineered human neural stem cells. The presence of the therapeutic genes CD and IFN-β was confirmed with reverse transcriptase–polymerase chain reaction (RT-PCR). After RNA extractions from both HB1.F3.CD and HB1.F3.CD.IFN-β cell lines, cDNAs were synthesized. cDNAs of the two cell lines were amplified through polymerase chain reaction (PCR) and the PCR products were confirmed with gel electrophoresis. The expressions of E. coli CD gene (472 bp) and human interferon-β gene (296 bp) were detected by RT-PCR. Glyceraldehyde 3-phosphate dehydrogenase gene (351 bp) was used as an internal control.
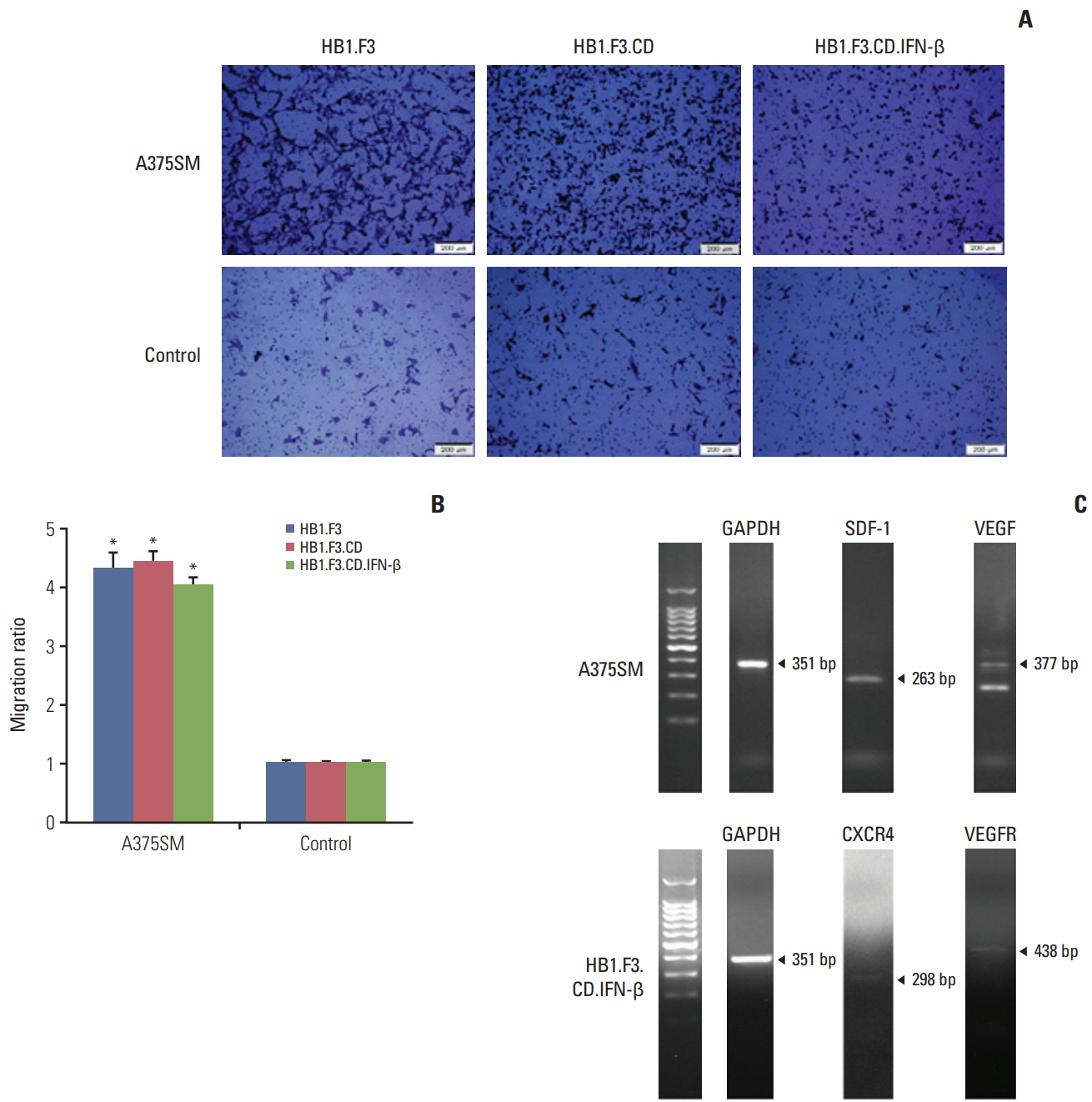
Fig. 2. Migratory effect of engineered human neural stem cells (hNSCs) with tumor tropic characteristics in a melanoma cell line (A375SM). (A) Normal human lung cells (L-132) and melanoma cells (A375SM) (1×105 cells/well) were seeded on a 24-well plate. hNSCs (1×105 cells/well) were seeded in the fibronectin pre-coated upper compartment of modified Boyden chambers. To observe the migrated hNSCs, crystal violet staining solution was added to both lower and upper compartments. (B) The migratory ratio of hNSCs was quantified using the ‘Cell Sense Dimension’ system (Olympus, Tokyo, Japan). Data were represented as mean±standard error of mean. *p < 0.05 vs. control. (C) Expression of various chemoattractant factors were displayed in A375SM cells. GAPDH, glyceraldehyde 3-phosphate dehydrogenase; SDF-1, stromal cell-derived factor 1; VEGF, vascular endothelial growth factor; VEGFR, vascular endothelial growth factor receptor.
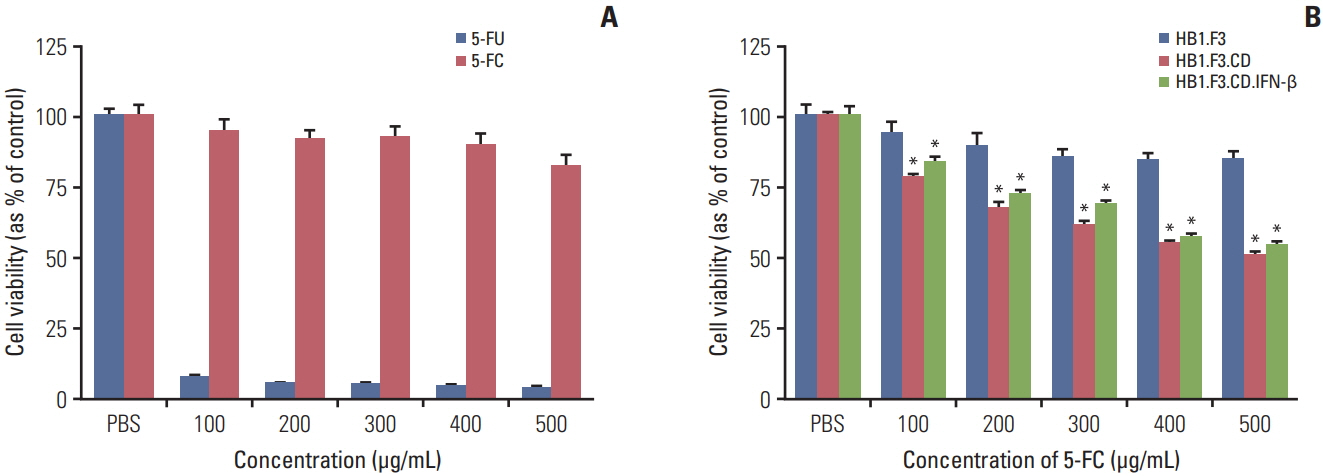
Fig. 3. Therapeutic efficacy of HB1.F3.CD and HB1.F3.CD.IFN-β with prodrug 5-fluorocytosine (5-FC) in vitro. Twenty-four hours after A375SM cells (4×103 cells/well) were seeded in 96-well plates, human neural stem cells (hNSCs) were seeded in the same well and all wells were treated with 5-FC for 3 days. The viability of the A375SM cells was measured using an MTT assay. (A) The cell viability of various concentrations of 5-FC and 5-fluorouracil (5-FU) on A375SM cells. (B) The therapeutic effect of hNSCs with 5-FC treatment at increasing concentrations (100, 200, 300, 400, and 500 μg/mL) on the A375SM co-culture system. Phosphate buffered saline (PBS) was used as a negative control. Data were represented as mean±standard error of mean. *p < 0.05 vs. control.
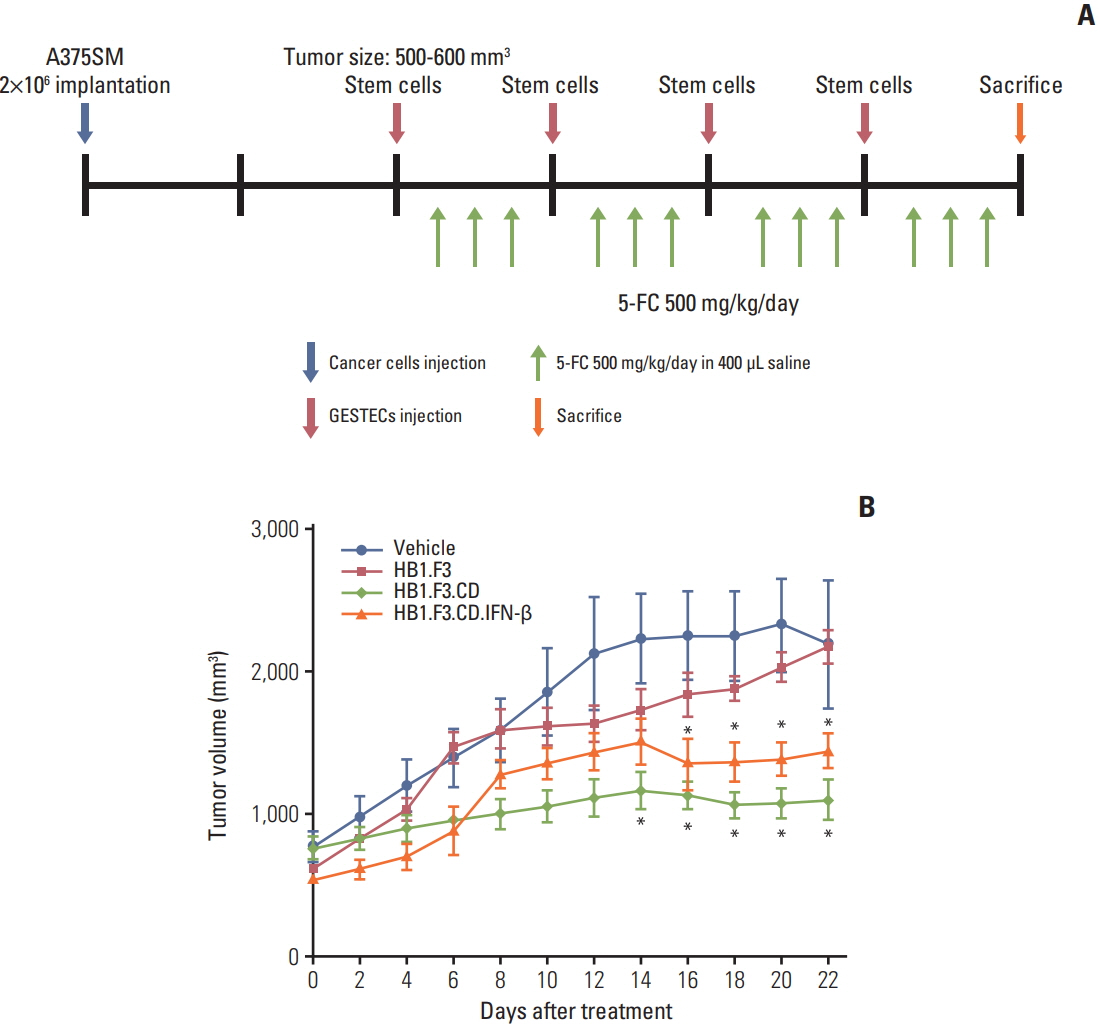
Fig. 4. Changes in tumor volume of melanoma A375SM cell line following human neural stem cell (hNSC) treatments. A xenograft model was created by implanting A375SM cells (2×106 cells) into female athymic mice. (A) CM-DiI pre-stained hNSCs (4×106 cells) were injected to adjacent tumor masses after the tumor mass reached between 500 and 600 mm3 . Intraperitoneal injection of 5-fluorocytosine (5-FC; 500 mg/kg/day) was implemented for 3 days after 2 days of hNSC injection. (B) The measurement of tumor volumes was carried out for 3 weeks and calculated as length×width×height×0.5236 (mm3 ). The changes in tumor after treatment with hNSCs in the presence of 5-FC were displayed in a graph. Data were represented as mean±standard error of mean. *p < 0.05 vs. 5-FC treatment alone.

Fig. 5. Fluorescence analysis of melanoma tumor mass in a xenograft mouse model. Human neural stem cells (hNSCs) were pre-labeled using a CM-DiI cell tracker. CM-DiI pre-labled hNSCs were injected during the experiment. DAPI counterstaining was conducted after the tumor section was prepared. The stained sections were able to be seen using fluorescence microscopy. ImageJ was used for merging the picture of DAPI and CM-DiI. Blue indicates DAPI stained nuclei of A375SM cells. Red and yellow arrows indicate CM-DiI pre-labled hNSCs.
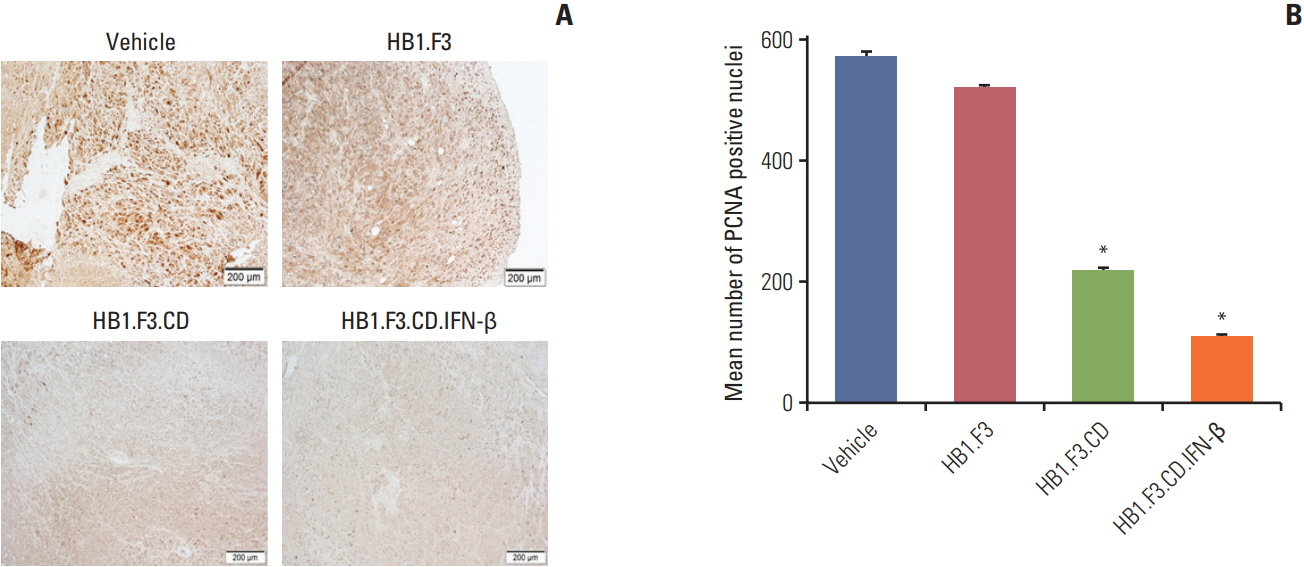
Fig. 6. Immunohistochemistry analysis of proliferating cell nuclear antigen (PCNA) in a tumor mass in melanoma xenograft mouse model. The expression of PCNA in tumor masses after immunohistochemical analysis was shown. Tumor masses were taken from every mouse. A 10% formalin fixation, paraffin embedding, and 5-μm sectioning using a microtome were performed. (A) A primary antibody specific for PCNA was used and the data were analyzed using a microscope. (B) The mean number of PCNA positive nuclei of each section was quantified and shown in the graph. Data are represented as mean±standard error of mean. *p < 0.05 vs. vehicle (5-fluorocytosine).
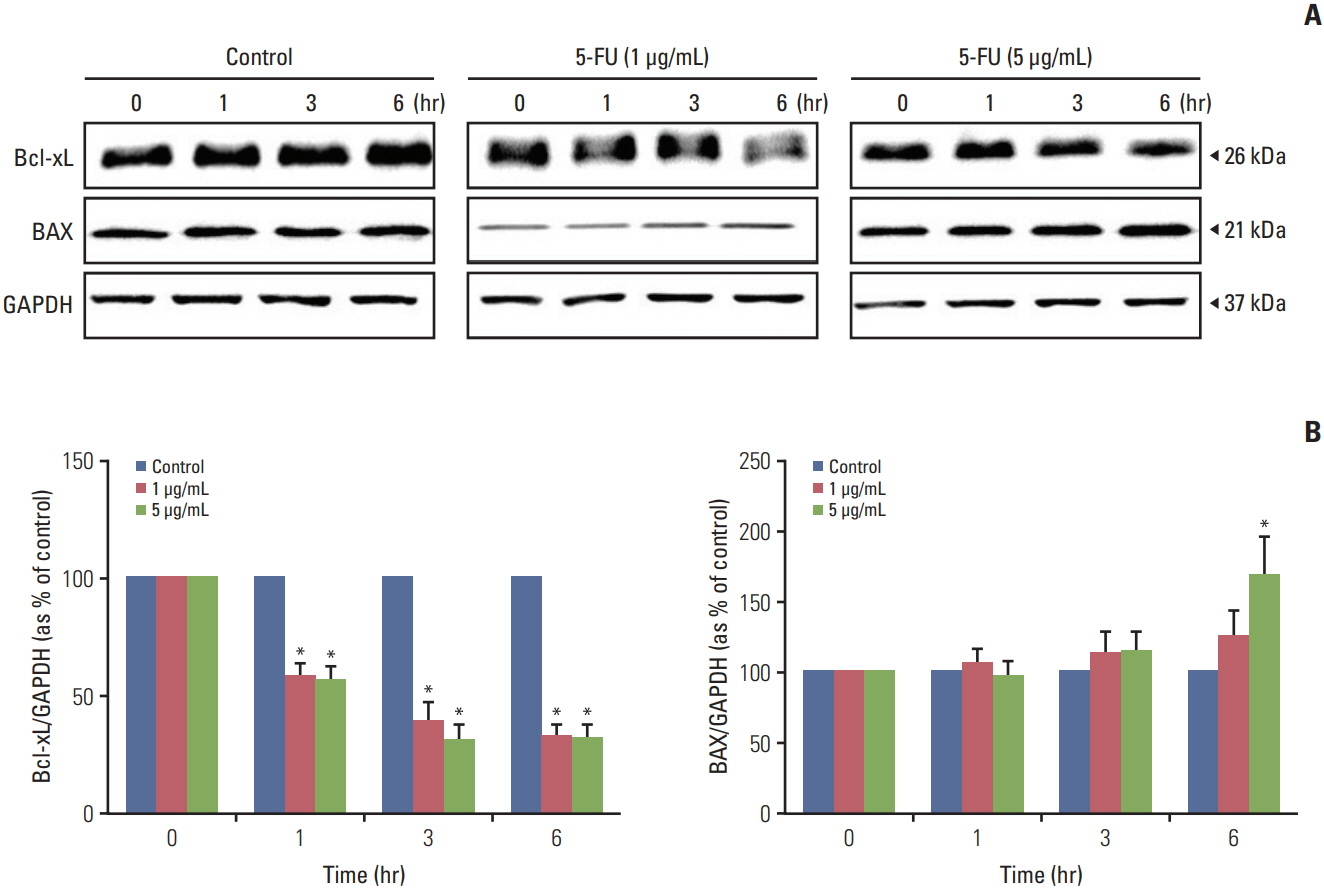
Fig. 7. Alteration of apoptosis related protein expression of melanoma cell line (A375SM) after 5-fluorouracil (5-FU) treatment. To evaluate the effects of 5-FU, diluted 5-FU (1 and 5 μg/mL) was applied to A375SM cells. Whole cell lysates were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis then immunoblotted with specific antibodies to Bcl-xL, BAX, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH). (A) Expression of Bcl-xL, BAX, and GAPDH proteins. (B) The relative value of Bcl-xL and BAX protein. Each protein level was normalized against GAPDH expression. Control, no treatment with 5-FU. Data are represented as the mean±standard deviation of three individual experiments each performed in three times. *p < 0.05 vs. control.
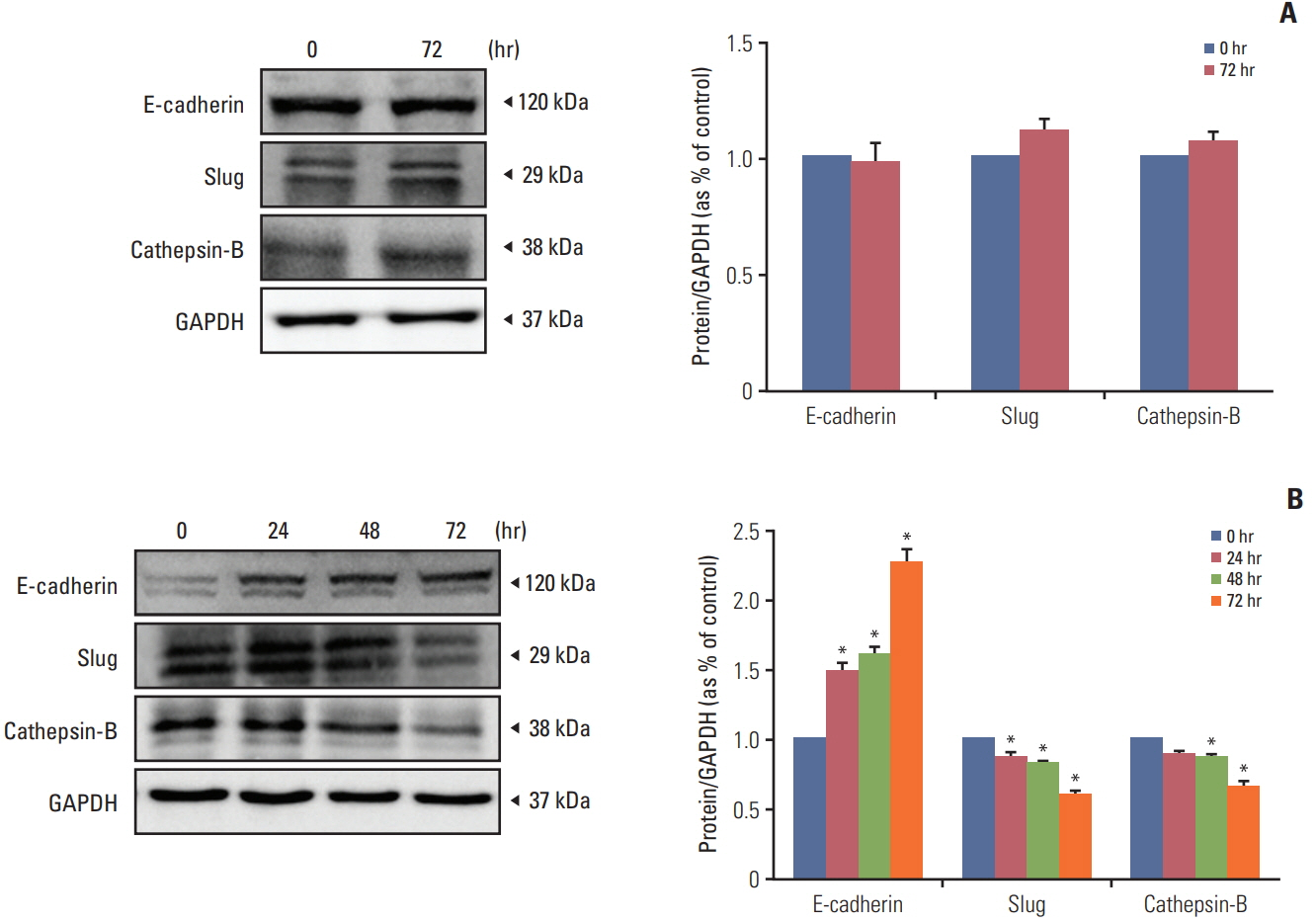
Fig. 8. Human interferon β from HB1.F3.CD.IFN-β cell line prevented epithelial-mesenchymal transition (EMT) and metastasis in melanoma cell line (A375SM). To confirm the effect of recombinant interferon-β of HB1.F3.CD.IFN-β on EMT and metastasis, the melanoma cell line (A375SM) was cultured with conditioned media from HB1.F3.CD.IFN-β for 0, 24, 48, and 72 hours. Expressions of epithelial protein E-cadherin, mesenchymal protein Slug, and metastasis protein cathepsin-B were measured by western blot. (A) The melanoma cell line was cultured with conditioned media from the HB1.F3.CD cell line as a control. (B) The melanoma cell line was cultured with conditioned media from HB1.F3.CD.IFN-β cell line. Data are displayed as the mean±standard deviation of three distinct experiments each conducted in triplicate. *p < 0.05 vs. 0 hour. GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Reference
-
References
1. American Cancer Society. Cancer facts and figures 2017. Atlanta, GA: American Cancer Society;2017.2. Berwick M, Buller DB, Cust A, Gallagher R, Lee TK, Meyskens F, et al. Melanoma epidemiology and prevention. Cancer Treat Res. 2016; 167:17–49.
Article3. Russak JE, Rigel DS. Risk factors for the development of primary cutaneous melanoma. Dermatol Clin. 2012; 30:363–8.
Article4. Marzagalli M, Montagnani Marelli M, Casati L, Fontana F, Moretti RM, Limonta P. Estrogen receptor beta in melanoma: from molecular insights to potential clinical utility. Front Endocrinol (Lausanne). 2016; 7:140.
Article5. Han J, Shi C, Dong X, Wang J, Wen H, Wang B, et al. Laparoscopic abdomino-perineal resection for patients with anorectal malignant melanoma: a report of 4 cases. J Biomed Res. 2016; 30:436–40.
Article6. Wellbrock C, Arozarena I. The complexity of the ERK/MAPkinase pathway and the treatment of melanoma skin cancer. Front Cell Dev Biol. 2016; 4:33.
Article7. Flaherty KT, McArthur G. BRAF, a target in melanoma: implications for solid tumor drug development. Cancer. 2010; 116:4902–13.8. Funck-Brentano E, Helias-Rodzewicz Z, Longvert C, Mokhtari K, Saiag P, Emile JF. Increase in NRAS mutant allele percentage during metastatic melanoma progression. Exp Dermatol. 2016; 25:472–4.9. Kim T, Amaria RN, Spencer C, Reuben A, Cooper ZA, Wargo JA. Combining targeted therapy and immune checkpoint inhibitors in the treatment of metastatic melanoma. Cancer Biol Med. 2014; 11:237–46.10. Yi BR, Park MA, Lee HR, Kang NH, Choi KJ, Kim SU, et al. Suppression of the growth of human colorectal cancer cells by therapeutic stem cells expressing cytosine deaminase and interferon-beta via their tumor-tropic effect in cellular and xenograft mouse models. Mol Oncol. 2013; 7:543–54.11. Yi BR, Hwang KA, Kang NH, Kim SU, Jeung EB, Kim HC, et al. Synergistic effects of genetically engineered stem cells expressing cytosine deaminase and interferon-beta via their tumor tropism to selectively target human hepatocarcinoma cells. Cancer Gene Ther. 2012; 19:644–51.12. Hernandez-Alcoceba R, Sangro B, Prieto J. Gene therapy of liver cancer. World J Gastroenterol. 2006; 12:6085–97.
Article13. Yi BR, O SN, Kang NH, Hwang KA, Kim SU, Jeung EB, et al. Genetically engineered stem cells expressing cytosine deaminase and interferon-beta migrate to human lung cancer cells and have potentially therapeutic anti-tumor effects. Int J Oncol. 2011; 39:833–9.14. Garrison JI, Berens ME, Shapiro JR, Treasurywala S, Floyd-Smith G. Interferon-beta inhibits proliferation and progression through S phase of the cell cycle in five glioma cell lines. J Neurooncol. 1996; 30:213–23.
Article15. Ren C, Kumar S, Chanda D, Kallman L, Chen J, Mountz JD, et al. Cancer gene therapy using mesenchymal stem cells expressing interferon-beta in a mouse prostate cancer lung metastasis model. Gene Ther. 2008; 15:1446–53.16. Ito S, Natsume A, Shimato S, Ohno M, Kato T, Chansakul P, et al. Human neural stem cells transduced with IFN-beta and cytosine deaminase genes intensify bystander effect in experimental glioma. Cancer Gene Ther. 2010; 17:299–306.17. Yi BR, Kim SU, Choi KC. Co-treatment with therapeutic neural stem cells expressing carboxyl esterase and CPT-11 inhibit growth of primary and metastatic lung cancers in mice. Oncotarget. 2014; 5:12835–48.
Article18. Aboody KS, Najbauer J, Danks MK. Stem and progenitor cellmediated tumor selective gene therapy. Gene Ther. 2008; 15:739–52.
Article19. Imitola J, Raddassi K, Park KI, Mueller FJ, Nieto M, Teng YD, et al. Directed migration of neural stem cells to sites of CNS injury by the stromal cell-derived factor 1alpha/CXC chemokine receptor 4 pathway. Proc Natl Acad Sci U S A. 2004; 101:18117–22.20. Schmidt NO, Przylecki W, Yang W, Ziu M, Teng Y, Kim SU, et al. Brain tumor tropism of transplanted human neural stem cells is induced by vascular endothelial growth factor. Neoplasia. 2005; 7:623–9.
Article21. Park GT, Kim SU, Choi KC. Anti-proliferative effect of engineered neural stem cells expressing cytosine deaminase and interferon-beta against lymph node-derived metastatic colorectal adenocarcinoma in cellular and xenograft mouse models. Cancer Res Treat. 2017; 49:79–91.22. Yi BR, Hwang KA, Aboody KS, Jeung EB, Kim SU, Choi KC. Selective antitumor effect of neural stem cells expressing cytosine deaminase and interferon-beta against ductal breast cancer cells in cellular and xenograft models. Stem Cell Res. 2014; 12:36–48.
Article23. Meng WC, Pan Y, Zhao X. Epirubicin-gold nanoparticles suppress hepatocellular carcinoma xenograft growth in nude mice. J Biomed Res. 2015; 29:486–90.
Article24. Mullen CA, Coale MM, Lowe R, Blaese RM. Tumors expressing the cytosine deaminase suicide gene can be eliminated in vivo with 5-fluorocytosine and induce protective immunity to wild type tumor. Cancer Res. 1994; 54:1503–6.25. Heo JR, Kim NH, Cho J, Choi KC. Current treatments for advanced melanoma and introduction of a promising novel gene therapy for melanoma (Review). Oncol Rep. 2016; 36:1779–86.
Article26. Bredholt G, Mannelqvist M, Stefansson IM, Birkeland E, Bo TH, Oyan AM, et al. Tumor necrosis is an important hallmark of aggressive endometrial cancer and associates with hypoxia, angiogenesis and inflammation responses. Oncotarget. 2015; 6:39676–91.
Article27. Longley DB, Harkin DP, Johnston PG. 5-fluorouracil: mechanisms of action and clinical strategies. Nat Rev Cancer. 2003; 3:330–8.
Article28. Yi BR, Kim SU, Choi KC. Synergistic effect of therapeutic stem cells expressing cytosine deaminase and interferon-beta via apoptotic pathway in the metastatic mouse model of breast cancer. Oncotarget. 2016; 7:5985–99.
Article29. Yi BR, Kim SU, Choi KC. Additional effects of engineered stem cells expressing a therapeutic gene and interferon-beta in a xenograft mouse model of endometrial cancer. Int J Oncol. 2015; 47:171–8.30. Kudryavets YI, Bezdenezhnykh NO, Lykhova OO, Semesiuk NI, Vorontsova AL. The role of interferon as a modifier of epithelial-mesenchymal transition in tumor cells. Exp Oncol. 2011; 33:178–81.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Anti-proliferative Effect of Engineered Neural Stem Cells Expressing Cytosine Deaminase and Interferon-β against Lymph Node–Derived Metastatic Colorectal Adenocarcinoma in Cellular and Xenograft Mouse Models
- Targeting Orthotopic Glioma in Mice with Genetically Engineered Salmonella typhimurium
- Current Concepts of Stem Cell Therapy
- Combination Therapy for Gliomas Using Temozolomide and Interferon-Beta Secreting Human Bone Marrow Derived Mesenchymal Stem Cells
- Glutathione Dynamics in the Tumor Microenvironment: A Potential Target of Cancer Stem Cells and T Cells
