Aging Effects on Dendritic Cells after Total Body Irradiation in Mice
- Affiliations
-
- 1Department of Hematology-Oncology, Ajou University School of Medicine, Suwon, Korea. jspark65@ajou.ac.kr
- KMID: 2305197
- DOI: http://doi.org/10.5045/kjh.2007.42.3.224
Abstract
-
BACKGROUND: It is still obscure how dendritic cells (DCs) can orchestrate whole immune reactions according to the host age. We studied changes of murine splenic DCs after total body irradiation (TBI), with regards to age.
METHODS
Young (8~14 wk) and old (12~16 mo) C57Bl/6 mice were irradiated with a dose of 1,100 cGy and were assessed 6 h later for phenotypic and functional changes of the DCs. The mean fluorescence intensities and cytokine producing cell proportions were analyzed with the student's t-test.
RESULTS
Interleukin-12 (IL-12), interferon (IFN gamma) and tumor necrosis factor (TNF alpha) producing classical DCs (cDCs) were more numerous in the young untreated mice than in the old mice. However, the number of these cells decreased in the young mice and increased in the old mice after TBI. IL-12, IFN gamma and TNF alpha producing plasmacytoid DCs (pDCs) were more frequent in the old mice than in the young mice before TBI both mice showed an increased frequency of cells producing these cytokines after TBI. Overall, the highest numbers of cDCs and pDCs producing IL-12, IFN gamma and TNF alpha were present in the old mice after TBI. In both the cDC and pDC populations, the old mice had a higher frequency of IL-10+ cells prior to TBI. After irradiation, the young mice had a higher frequency of IL-10+ cells.
CONCLUSION
With TBI, the DCs showed dramatic differences between young and old mice. Young mice turned to an immuno-suppressive response whereas the old mice changed to an immuno-stimulation of DCs after TBI. From these dramatic aging effects, we hope to explain the different frequencies and severities of acute GvHD after allogeneic hematopoietic stem cell transplantation according to host age.
Keyword
MeSH Terms
Figure
-
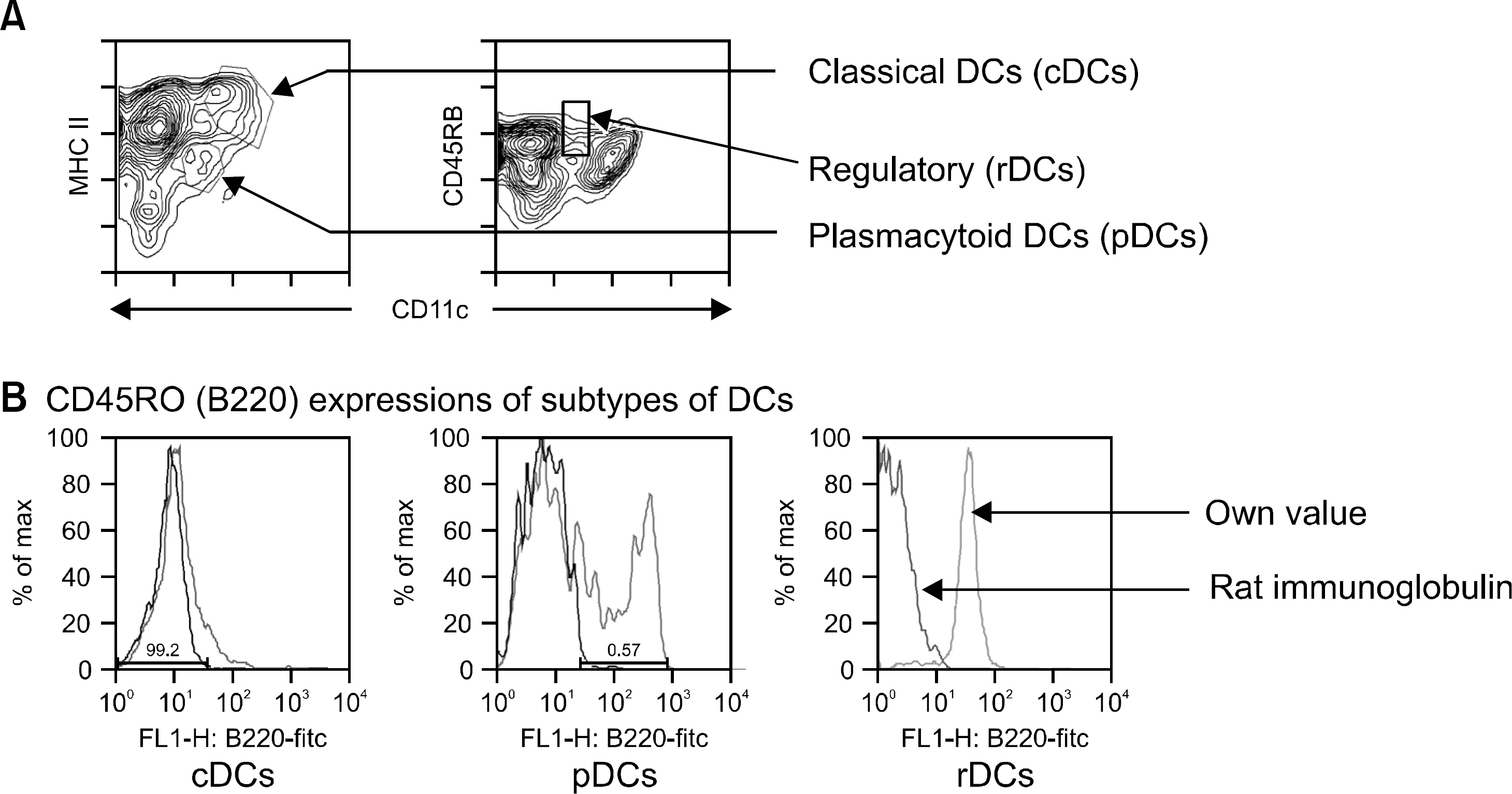
Fig. 1 Discrimination of subtypes of dendritic cells. (A) CD11chi, MHC class IIhi populations represented classical DDs (cDCs), CD11clo, MHC class IIlo populations were plasmacytoid DCs (pDCs), and CD11clo, CD45RBhi were regulatory DCs (rDCs). (B) Whereas cDCs failed to express B220, pDCs eapressed B220. rDCs showed B220 positive unlike other study (10). The hamster monoclonal antibody N418 reacts with an epitope of the β2 integrin family heterodimer p150/95 (CD11c).

Fig. 2 Changes of MFI of co-stimulatory molecules on DCs before versus after TBI or young versus old mice. While young mice did not show any change of co-stimulatory molecules on DCs after TBI, old mice were observed up-regulations of CD80, CD86, and CD40 of cDCs and pDCs. But resting levels of co-stimulatory molecules between young and old mice were not different each other, as was after TBI (data not shown). ∗represents P value<0.05 by two-tailed student t-test.

Fig. 3 MFI of IFN-γ before versus after TBI, young versus old mice. MFI of IFN-γ in young cDCs increased TBI (P=0.0201) but the values of rat immunoglobulin (negative control, †) was so high that the measured values could be distorted. Therefore, the MFI of IFN-γdid not show significant change sequentially (before vs. after TBI) or cross sectionally (young vs. old mice). ∗represents P value<0.05 by two-tailed student t-test.
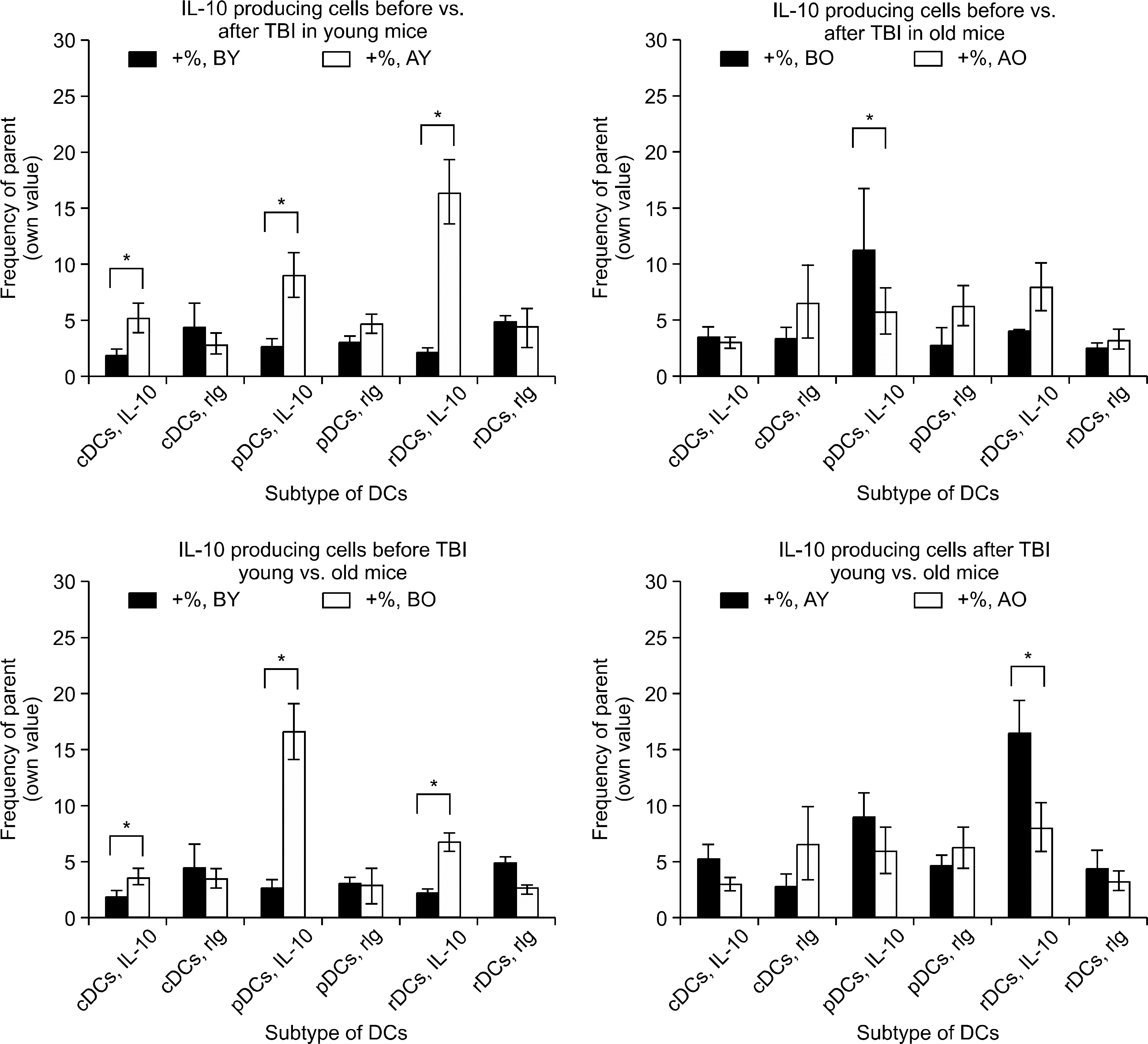
Fig. 4 Changes of IL-10 producing cells, before versus after TBI, young versus old mice. IL-10 producing cells in young mice significantly increased after TBI in all three subgroups of DCs (P=0.0375, 0.0051, and 0.0019 for cDCs, pDCs, and rDCs, respectively). However in old mice, TBI increased only pDCs of IL-10 producing cells (P=0.0149). Interestingly, at resting state, old mice had IL-10 producing cells much more than young mice (P=0.0357, 0.0027, and 0.0011 for cDCs, pDCs, and rDCs) but after TBI, the differences disappeared or reversed (P=0.0382 for rDCs). ∗represents P value<0.05 by two-tailed student t-test.
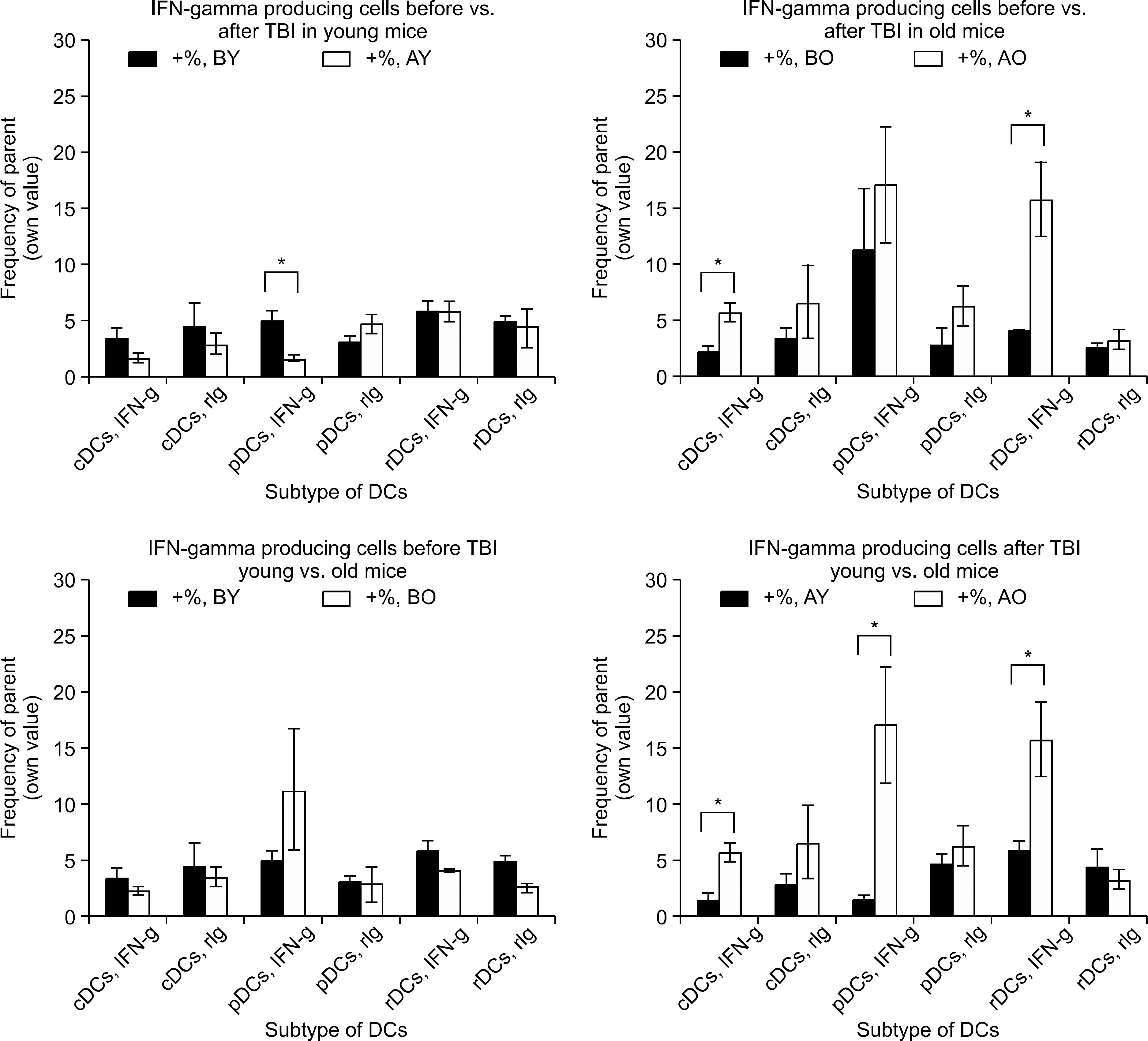
Fig. 5 Changes of IFN-γ producing cells, before versus after TBI, young versus old mice. Count of IFN-γ producing cells showed reversed phenomenon over IL-10. Young mice decreased (P=0.0087 for pDCs) or no change but old mice generally increased IFN-γ producing cells after TBI (P=0.0068, 0.0077 for cDCs, rDCs, respectively). Before TBI, young and old mice did not show difference of IFN-γ producing cell number however after TBI, IFN-γ producing cell number of all three subtypes of DCs were higher in old mice than in young mice (P=0.0041, 0.00132, and 0.0057 for cDCs, pDCs). ∗represents P value<0.05 by two-tailed student t-test.
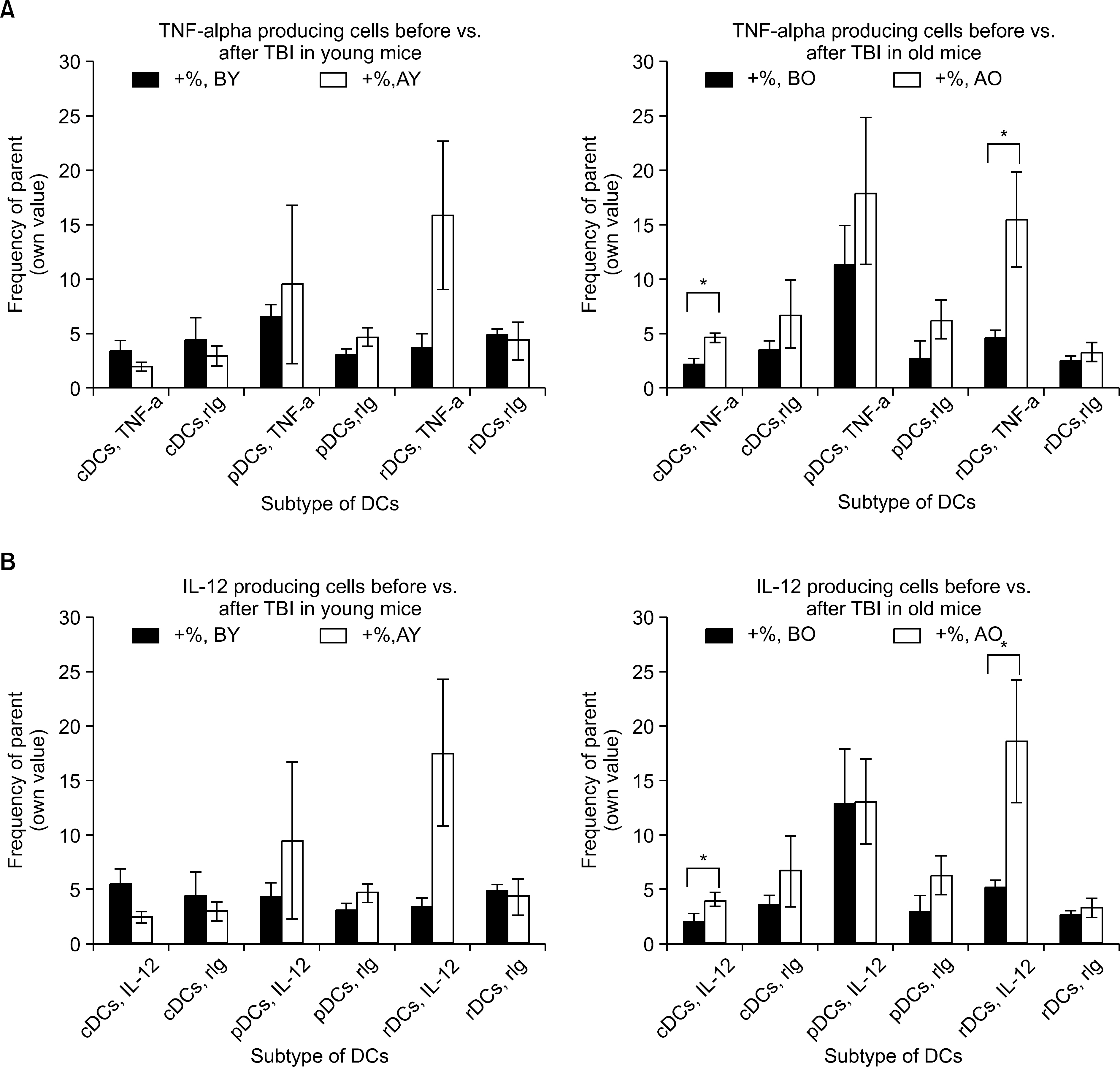
Fig. 6 Changes of TNF-α & IL-12 producing cells, before versus after TBI, young versus old mice. (A) TNF-α producing cells commonly increased in young and old mice after TBI, cDCs of old mice showed significant increment of the cell counts (P=0.008 and 0.0187 for cDCs and rDCs, respectively). Expect for higher numbers of TNF-α producing cells in old cDCs (P=0.0167), cross-sectional analysis failed to show any difference between young old mice. (B) IL-12 producing cells increased significantly in old mice after TBI, not in young mice (P=0.0094 and 0.0218 for cDCs and rDCs). Before TBI or after TBI there was no difference of IL-12 producing cell count between young and old mice. ∗represents P value<0.05 by two-tailed student t-test.
Reference
-
1). Maraninchi D., Gluckman E., Blaise D, et al. Impact of T-cell depletion on outcome of allogeneic bone-marrow transplantation for standard-risk leukae-mias. Lancet. 1987. 2:175–8.
Article2). Marmont AM., Horowitz MM., Gale RP, et al. T-cell depletion of HLA-identical transplants in leukemia. Blood. 1991. 78:2120–30.
Article3). Sullivan K., Storb R., Buckner CD, et al. Graft-versus-host disease as adoptive immunotherapy in patients with advanced hematologic neoplasms. N Engl J Med. 1989. 320:828–34.
Article4). Bryson JS., Jennings CD., Caywood BE., Dix AR., Lowery DM., Kaplan AM. Enhanced graft-versus-host disease in older recipient mice following allogeneic bone marrow transplantation. Bone Marrow Transplant. 1997. 19:721–8.
Article5). Storb R. Graft rejection and graft-versus-host disease in marrow transplantation. Transplant Proc. 1989. 21:2915–8.6). Shlomchik WD., Couzens MS., Tang CB, et al. Prevention of graft versus host disease by inactivation of host antigen-presenting cells. Science. 1999. 285:412–5.
Article7). Steinman RM., Cohn ZA. Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J Exp Med. 1973. 137:1142–62.8). Steinman RM., Lusting DS., Cohn ZA. Identification of a novel cell type in peripheral lympoid organs of mice. III. Functional properties in vivo. J Exp Med. 1974. 139:1431–45.9). Steinman RM. The dendritic cell system and its role in immunogenicity. Annu Rev Immunol. 1991. 9:271–96.
Article10). Wakkach A., Fournier N., Brun V., Breittmayer JP., Cottrez F., Groux H. Characterization of dendritic cells that induce tolerance and T regulatory 1 cell differentiation in vivo. Immunity. 2003. 18:605–17.
Article11). Sato K., Yamashita N., Baba M., Matsuyama T. Modified myeloid dendritic cells act as regulatory dendritic cells to induce anergic and regulatory T cells. Blood. 2003. 101:3581–9.
Article12). Sato K., Yamashita N., Yamashita N., Baba M., Matsuyama T. Regulatory dendritic cells protect mice from murine acute graft-versus-host disease and leukemia relapse. Immunity. 2003. 18:367–79.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Acute Response to Co-60 Total Body Irradiation (TBI) With 600 cgy at 3 Different Does Rates in the Mice
- Effects of X-irradiation on the levels of serum cholesterol in mouse
- Effects of same TDF Factors on Body Weight of Mice and Peripheral Blood Picture
- Defects in the differentiation and function of bone marrow-derived dendritic cells in non-obese diabetic mice
- Aspectual Comparison of the Skin Changes in Hairless Mice According to the Aging Type
