Effects of Interleukin-9 Blockade on Chronic Airway Inflammation in Murine Asthma Models
- Affiliations
-
- 1Department of Internal Medicine, Soonchunhyang University Gumi Hospital, Gumi, Korea.
- 2Department of Microbiology, Ewha Womans University School of Medicine, Seoul, Korea. soyounwoo@ewha.ac.kr
- 3Department of Internal Medicine, Ewha Womans University School of Medicine, Seoul, Korea.
- KMID: 2260312
- DOI: http://doi.org/10.4168/aair.2013.5.4.197
Abstract
- PURPOSE
Asthma is a chronic inflammatory disease of the airways associated with structural changes and airway remodeling. Interleukin (IL)-9 has pleiotropic effects on both inflammatory cells and airway structural cells, which are involved in asthma pathogenesis. We evaluated the effects of IL-9 blockade on chronic airway inflammation.
METHODS
Acute airway inflammation was induced in Balb/c mice using aerosolized ovalbumin (OVA), whereas chronic asthma was induced by OVA exposure for 5 weeks with anti-IL-9 or isotype-matched antibody (Ab) treatment during the OVA challenge. Inflammatory cells in bronchoalveolar lavage fluid (BALF) were counted and lung tissues were stained to detect cellular infiltration, mucus deposition, and collagen accumulation. The levels of interferon (IFN)-gamma, IL-4, IL-5, IL-9, IL-17, and immunoglobulin E (IgE) in BALF were measured using enzyme linked immunosorbent assays, and profiles of inflammatory cells and subsets of T helper (Th) cells were analyzed using flow cytometry.
RESULTS
IL-9, IL-17, and IFN-gamma levels were significantly increased in the chronic group compared to the acute asthma group. However, the number of IL-9-positive cells was not affected, with a decrease in Th17 cells in OVA-challenged caspase-1 knockout mice. Numbers of eosinophils, neutrophils, B cells, mast cells, and Th17 cells decreased after administration of anti-IL-9 Ab. Total IgE, IL-5, IL-9, and IL-17 levels were also lower in the anti-IL-9 group.
CONCLUSIONS
Our results suggest that anti-IL-9 Ab treatment inhibits pulmonary infiltration of inflammatory cells and cytokine production, especially IL-17. These results provide a basis for the use of an anti-IL-9 Ab to combat IL-17-mediated airway inflammation.
MeSH Terms
-
Airway Remodeling
Animals
Asthma
B-Lymphocytes
Bronchoalveolar Lavage Fluid
Collagen
Eosinophils
Immunoglobulin E
Immunoglobulins
Inflammation
Interferons
Interleukin-17
Interleukin-4
Interleukin-5
Interleukin-9
Interleukins
Lung
Mast Cells
Mice
Mice, Knockout
Mucus
Neutrophils
Ovalbumin
Ovum
Th17 Cells
Collagen
Immunoglobulin E
Immunoglobulins
Interferons
Interleukin-17
Interleukin-4
Interleukin-5
Interleukin-9
Interleukins
Ovalbumin
Figure
-

Fig. 1 Protocol for inducing experimental acute and chronic airway inflammation models. (A) In the acute asthma model, mice were sensitized by two intraperitoneal injections of OVA on days 0 and 5, and then received aerosolized OVA challenge on days 12-15 using a nebulizer. (B) Mice were sensitized via intraperitoneal injection of OVA on days 0 and 12 to induce chronic asthma. Mice were exposed to an aerosol of 1% OVA in PBS on days 18-23 using a nebulizer. After day 26, mice were exposed to aerosolized 1% OVA three times per week for 5 weeks. OVA, ovalbumin; PBS, phosphate-buffered saline.
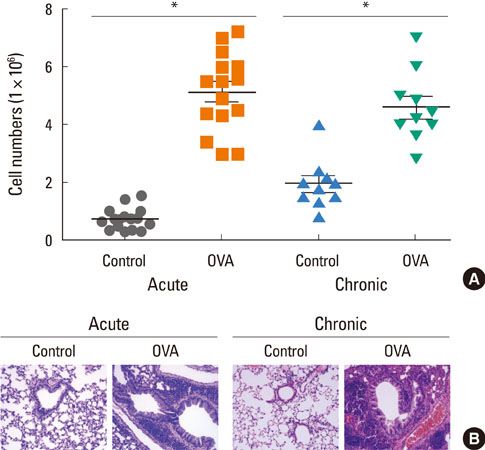
Fig. 2 OVA-induced airway inflammation in acute and chronic asthma groups. (A) BALF from acute (n=15) or chronic asthma (n=10) mice was collected 24 hours after the final OVA challenge, and the number of cells in the BALF was determined as described in the Materials and Methods. The data are expressed as the means±SEM (*P<0.05, groups were compared as indicated by Mann-Whitney U tests). (B) Mice were sacrificed 1 day after the last OVA challenge. Mice lungs were fixed with 4% paraformaldehyde, and tissue sections were stained with H&E. Images are presented at the original magnification of ×400. OVA, ovalbumin; BALF, bronchoalveolar lavage fluid; SEM, standard error of the mean; H&E, hematoxylin and eosin.
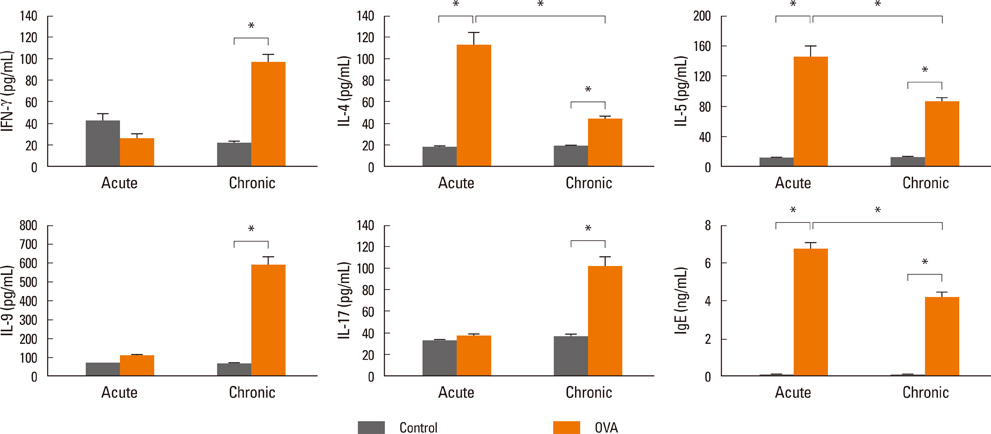
Fig. 3 Comparison of cytokine profiles after OVA challenge. Concentrations of IFN-γ, IL-4, IL-5, IL-9, IL-17, and IgE in BALF from acute asthma or chronic asthma mice were measured by ELISA. Data are expressed as the means±SEM (n=5). *P<0.05 for control versus OVA-inhalation and acute-OVA versus chronic-OVA by Mann-Whitney U tests. OVA, ovalbumin; IFN, interferon; IL, interleukin; BALF, bronchoalveolar lavage fluid; ELISA, enzyme-linked immunosorbent assay; SEM, standard error of the mean.
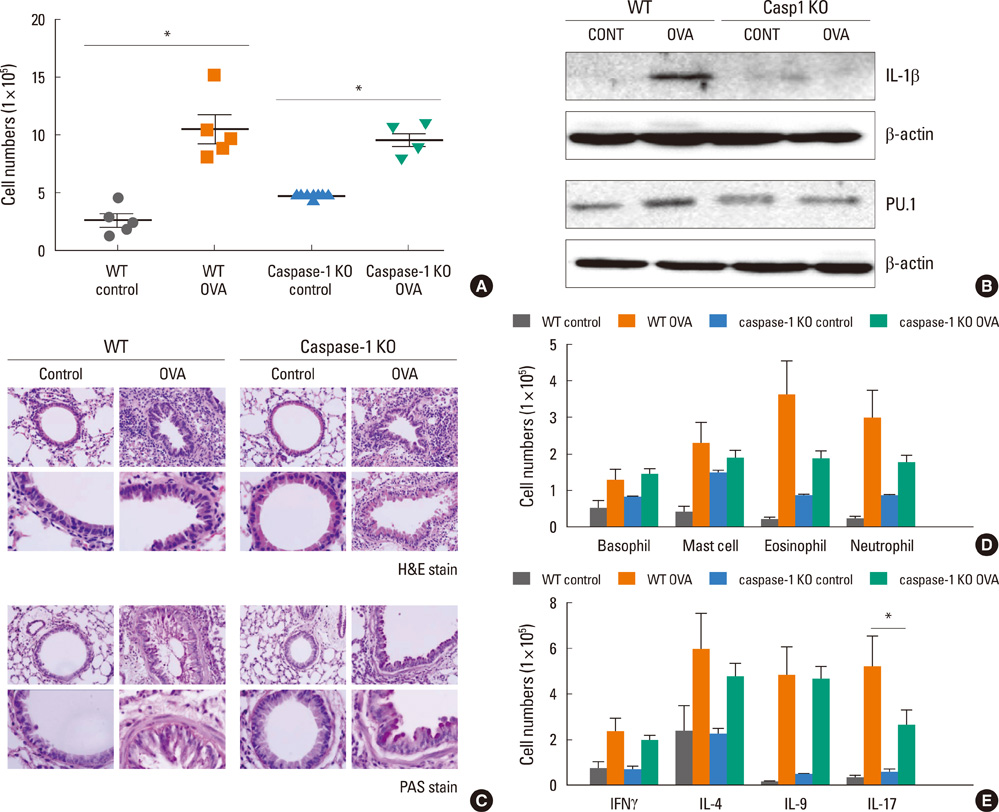
Fig. 4 Caspase-1 KO mice induced airway inflammation. (A) BALF from WT C57BL/6 (n=5) or caspase-1 KO (n=5) mice was collected 24 hours after the final OVA challenge, and the number of cells in the BALF was determined. The data are expressed as the means±SEM (*P<0.05;0.05, groups were compared as indicated by Mann-Whitney U tests). (B) BAL cells were lysed, and whole-cell extracts were resolved using 12% SDS-PAGE, transferred to polyvinylidene fluoride membranes, and probed with anti-IL-1β, anti-β-actin, and anti-PU.1 Ab. Representative blots of three independent experiments are shown. (C) Mice lungs were fixed with 4% paraformaldehyde, tissue sections were stained with H&E, and mucus was visualized by PAS staining. Images are presented at the original magnification of ×400. (D) After OVA challenge, proportions of basophils (CD49d+, FcεRI+), mast cells (c-Kit+, SSClow), eosinophils (CD49b+, SSCint), and neutrophils (Gr1+) were collected for flow cytometric analysis (n=5 for each groups). Cell numbers were calculated from the percentage of cells×BAL cell numbers. (E) The proportions of CD4+ T cells were determined according to intracellular levels of cytokines: IFN-γ+ for IFN-γ-secreting T cells, IL-4+ for IL-4-secreting T cells, IL-9+ for IL-9-secreting T cells, and IL-17+ for Th17 cells. Cell numbers were calculated from the percentage of cells subsets×BAL cell numbers. Data are expressed as the means±SEM (n=5). *P<0.05; groups were compared as indicated by unpaired t-tests. KO, knockout; BALF, bronchoalveolar lavage fluid; OVA, ovalbumin; SEM, standard error of the mean; SDS-PAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis; IL, interleukin; Ab, antibody; H&E, hematoxylin and eosin; PAS, periodic acid-Schiff.

Fig. 5 Effects of IL-9 blockade on chronic pulmonary inflammation. (A) Anti-IL-9 or control Iso IgG Ab were administered (100 µg/mouse, intraperitoneally) once a week throughout the OVA induced development of chronic asthma. Five weeks of OVA challenge induced massive infiltration of eosinophils, and anti-IL-9 treatment significantly suppressed their infiltration. (B) BALF from each experimental mouse was collected 24 hours after the final OVA challenge, and the number of cells in the BALF was determined. (C) Differential cell counts were calculated as (total cell number of BAL cells)×(percentage of mononuclear cells, eosinophils, or neutrophils counted in the Giemsa-stained cytospin cell preparation). MNC=mononuclear cell. (D) IL-9 blockade attenuated tissue inflammation. The randomly selected histologic images were scored as the mean inflammatory index, which represents the severity of peri-bronchial inflammation. Mucus producing goblet cells were visualized by PAS staining. Sections were stained with Sirius Red solution to detect collagen deposition. Images are presented at the original magnification of ×400. PAS-positive goblet cells and Sirius Red-positive sub-epithelial areas for each bronchiole were calculated using image analysis software for quantitative analysis. Data are expressed as the mean±SEM (n=5). *P<0.05; groups were compared as indicated using the Mann-Whitney U tests. OVA, ovalbumin; BALF, bronchoalveolar lavage fluid; IL, interleukin; PAS, periodic acid-Schiff; SEM, standard error of the mean.
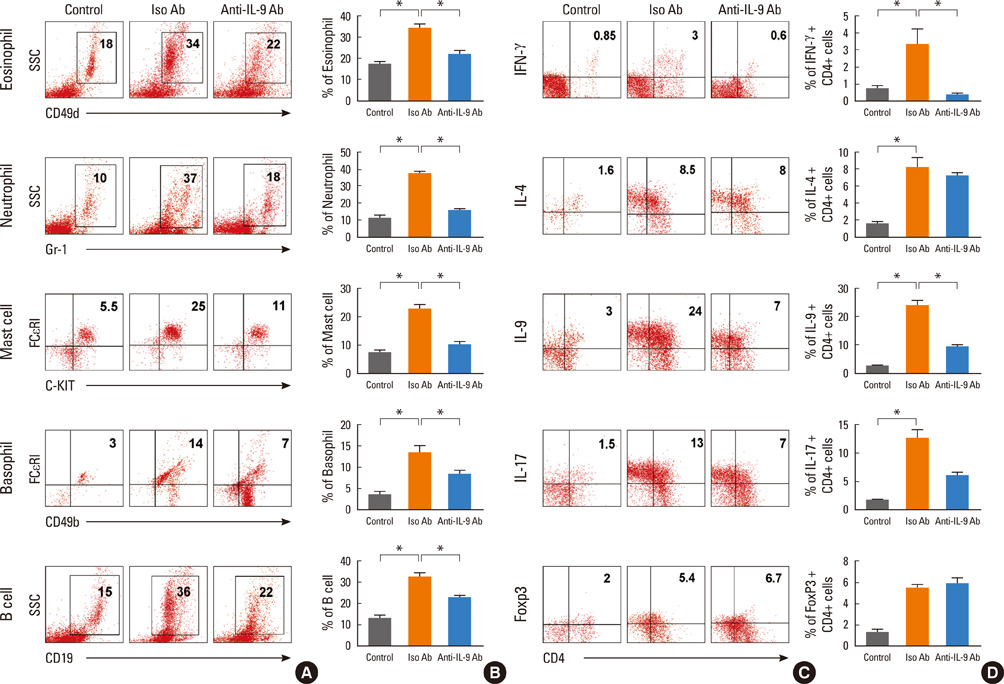
Fig. 6 IL-9 blockade altered BAL cell profiles. Flow cytometric analysis shows profiles of cells from BALF. Eosinophils (CD49b+, SSCint), neutrophils (Gr1+), mast cells (c-Kit+, SSClow), basophils (CD49d+, FcεRI+), and B cells (CD19+) were assessed. Data are presented as representative results of three independent experiments (A) and a statistical graph (B). The proportions of CD4+ T cells were determined according to intracellular levels of cytokines: IFN-γ+ for IFN-γ-secreting T cells, IL-4+ for IL-4-secreting T cells, IL-9+ for IL-9-secreting T cells, IL-17+ for Th17 cells, and Foxp3+ for Treg cells. Data are presented as representative results of three independent experiments (C) and a statistical graph (D). Data are expressed as the means±SEM (n=5) *P<0.05; groups were compared as indicated by unpaired t-tests. BALF, bronchoalveolar lavage fluid; SEM, standard error of the mean.

Fig. 7 IL-9 blockade alters cytokine levels in BALF. Mice were sacrificed and the levels of IgE and cytokines in BALF from OVA-challenged mice with anti-IL-9 Ab or Iso IgG Ab were measured using ELISA. Data are expressed as the means±SEM (n=5). *P<0.05; groups were compared as indicated by Mann-Whitney U tests. IL, interleukin; BALF, bronchoalveolar lavage fluid; OVA, ovalbumin; ELISA, enzyme-linked immunosorbent assay; SEM, standard error of the mean.
Cited by 2 articles
-
Magnolol exerts anti-asthmatic effects by regulating Janus kinase-signal transduction and activation of transcription and Notch signaling pathways and modulating Th1/Th2/Th17 cytokines in ovalbumin-sensitized asthmatic mice
Qi Huang, Lele Han, Rong Lv, Ling Ling
Korean J Physiol Pharmacol. 2019;23(4):251-261. doi: 10.4196/kjpp.2019.23.4.251.Anti-Interleukin-9 Antibody Increases the Effect of Allergen-Specific Immunotherapy in Murine Allergic Rhinitis
Ji-Hyeon Shin, Do Hyun Kim, Boo-Young Kim, Sung Won Kim, Se Hwan Hwang, Joohyung Lee, Soo Whan Kim
Allergy Asthma Immunol Res. 2017;9(3):237-246. doi: 10.4168/aair.2017.9.3.237.
Reference
-
1. Global Initiative for Asthma (GINA). Global Strategy for Asthma Management and Prevention [Internet]. Vancouver (WA): GINA;2011. Available from: http://www.ginasthma.org/.2. Yamauchi K, Inoue H. Airway remodeling in asthma and irreversible airflow limitation-ECM deposition in airway and possible therapy for remodeling. Allergol Int. 2007; 56:321–329.3. Yamauchi K. Airway remodeling in asthma and its influence on clinical pathophysiology. Tohoku J Exp Med. 2006; 209:75–87.4. Durrani SR, Viswanathan RK, Busse WW. What effect does asthma treatment have on airway remodeling? Current perspectives. J Allergy Clin Immunol. 2011; 128:439–448.5. Payne DN, Rogers AV, Adelroth E, Bandi V, Guntupalli KK, Bush A, Jeffery PK. Early thickening of the reticular basement membrane in children with difficult asthma. Am J Respir Crit Care Med. 2003; 167:78–82.6. Cho KA, Suh JW, Sohn JH, Park JW, Lee H, Kang JL, Woo SY, Cho YJ. IL-33 induces Th17-mediated airway inflammation via mast cells in ovalbumin-challenged mice. Am J Physiol Lung Cell Mol Physiol. 2012; 302:L429–L440.7. Buc M, Dzurilla M, Vrlik M, Bucova M. Immunopathogenesis of bronchial asthma. Arch Immunol Ther Exp (Warsz). 2009; 57:331–344.8. Kaiko GE, Foster PS. New insights into the generation of Th2 immunity and potential therapeutic targets for the treatment of asthma. Curr Opin Allergy Clin Immunol. 2011; 11:39–45.9. Nair P, Pizzichini MM, Kjarsgaard M, Inman MD, Efthimiadis A, Pizzichini E, Hargreave FE, O'Byrne PM. Mepolizumab for prednisone-dependent asthma with sputum eosinophilia. N Engl J Med. 2009; 360:985–993.10. Haldar P, Brightling CE, Hargadon B, Gupta S, Monteiro W, Sousa A, Marshall RP, Bradding P, Green RH, Wardlaw AJ, Pavord ID. Mepolizumab and exacerbations of refractory eosinophilic asthma. N Engl J Med. 2009; 360:973–984.11. Corren J. Cytokine inhibition in severe asthma: current knowledge and future directions. Curr Opin Pulm Med. 2011; 17:29–33.12. Berry MA, Hargadon B, Shelley M, Parker D, Shaw DE, Green RH, Bradding P, Brightling CE, Wardlaw AJ, Pavord ID. Evidence of a role of tumor necrosis factor alpha in refractory asthma. N Engl J Med. 2006; 354:697–708.13. Goswami R, Kaplan MH. A brief history of IL-9. J Immunol. 2011; 186:3283–3288.14. Chang HC, Sehra S, Goswami R, Yao W, Yu Q, Stritesky GL, Jabeen R, McKinley C, Ahyi AN, Han L, Nguyen ET, Robertson MJ, Perumal NB, Tepper RS, Nutt SL, Kaplan MH. The transcription factor PU.1 is required for the development of IL-9-producing T cells and allergic inflammation. Nat Immunol. 2010; 11:527–534.15. Putheti P, Awasthi A, Popoola J, Gao W, Strom TB. Human CD4 memory T cells can become CD4+IL-9+ T cells. PLoS One. 2010; 5:e8706.16. Staudt V, Bothur E, Klein M, Lingnau K, Reuter S, Grebe N, Gerlitzki B, Hoffmann M, Ulges A, Taube C, Dehzad N, Becker M, Stassen M, Steinborn A, Lohoff M, Schild H, Schmitt E, Bopp T. Interferon-regulatory factor 4 is essential for the developmental program of T helper 9 cells. Immunity. 2010; 33:192–202.17. McLane MP, Haczku A, van de Rijn M, Weiss C, Ferrante V, MacDonald D, Renauld JC, Nicolaides NC, Holroyd KJ, Levitt RC. Interleukin-9 promotes allergen-induced eosinophilic inflammation and airway hyperresponsiveness in transgenic mice. Am J Respir Cell Mol Biol. 1998; 19:713–720.18. Postma DS, Bleecker ER, Amelung PJ, Holroyd KJ, Xu J, Panhuysen CI, Meyers DA, Levitt RC. Genetic susceptibility to asthma--bronchial hyperresponsiveness coinherited with a major gene for atopy. N Engl J Med. 1995; 333:894–900.19. Nicolaides NC, Holroyd KJ, Ewart SL, Eleff SM, Kiser MB, Dragwa CR, Sullivan CD, Grasso L, Zhang LY, Messler CJ, Zhou T, Kleeberger SR, Buetow KH, Levitt RC. Interleukin 9: a candidate gene for asthma. Proc Natl Acad Sci U S A. 1997; 94:13175–13180.20. Choi JM, Ahn MH, Chae WJ, Jung YG, Park JC, Song HM, Kim YE, Shin JA, Park CS, Park JW, Park TK, Lee JH, Seo BF, Kim KD, Kim ES, Lee DH, Lee SK, Lee SK. Intranasal delivery of the cytoplasmic domain of CTLA-4 using a novel protein transduction domain prevents allergic inflammation. Nat Med. 2006; 12:574–579.21. Sutton C, Brereton C, Keogh B, Mills KH, Lavelle EC. A crucial role for interleukin (IL)-1 in the induction of IL-17-producing T cells that mediate autoimmune encephalomyelitis. J Exp Med. 2006; 203:1685–1691.22. Bullens DM, Truyen E, Coteur L, Dilissen E, Hellings PW, Dupont LJ, Ceuppens JL. IL-17 mRNA in sputum of asthmatic patients: linking T cell driven inflammation and granulocytic influx? Respir Res. 2006; 7:135.23. Erpenbeck VJ, Hohlfeld JM, Discher M, Krentel H, Hagenberg A, Braun A, Krug N. Increased expression of interleukin-9 messenger RNA after segmental allergen challenge in allergic asthmatics. Chest. 2003; 123:370S.24. Shimbara A, Christodoulopoulos P, Soussi-Gounni A, Olivenstein R, Nakamura Y, Levitt RC, Nicolaides NC, Holroyd KJ, Tsicopoulos A, Lafitte JJ, Wallaert B, Hamid QA. IL-9 and its receptor in allergic and nonallergic lung disease: increased expression in asthma. J Allergy Clin Immunol. 2000; 105:108–115.25. Bhathena PR, Comhair SA, Holroyd KJ, Erzurum SC. Interleukin-9 receptor expression in asthmatic airways In vivo. Lung. 2000; 178:149–160.26. Abdelilah S, Latifa K, Esra N, Cameron L, Bouchaib L, Nicolaides N, Levitt R, Hamid Q. Functional expression of IL-9 receptor by human neutrophils from asthmatic donors: role in IL-8 release. J Immunol. 2001; 166:2768–2774.27. Temann UA, Ray P, Flavell RA. Pulmonary overexpression of IL-9 induces Th2 cytokine expression, leading to immune pathology. J Clin Invest. 2002; 109:29–39.28. Townsend JM, Fallon GP, Matthews JD, Smith P, Jolin EH, McKenzie NA. IL-9-deficient mice establish fundamental roles for IL-9 in pulmonary mastocytosis and goblet cell hyperplasia but not T cell development. Immunity. 2000; 13:573–583.29. Cheng G, Arima M, Honda K, Hirata H, Eda F, Yoshida N, Fukushima F, Ishii Y, Fukuda T. Anti-interleukin-9 antibody treatment inhibits airway inflammation and hyperreactivity in mouse asthma model. Am J Respir Crit Care Med. 2002; 166:409–416.30. Kearley J, Erjefalt JS, Andersson C, Benjamin E, Jones CP, Robichaud A, Pegorier S, Brewah Y, Burwell TJ, Bjermer L, Kiener PA, Kolbeck R, Lloyd CM, Coyle AJ, Humbles AA. IL-9 governs allergen-induced mast cell numbers in the lung and chronic remodeling of the airways. Am J Respir Crit Care Med. 2011; 183:865–875.31. Doherty TA, Soroosh P, Broide DH, Croft M. CD4+ cells are required for chronic eosinophilic lung inflammation but not airway remodeling. Am J Physiol Lung Cell Mol Physiol. 2009; 296:L229–L235.32. Soroosh P, Doherty TA. Th9 and allergic disease. Immunology. 2009; 127:450–458.33. Parker JM, Oh CK, La Force C, Miller SD, Pearlman DS, Le C, Robbie GJ, White WI, White B, Molfino NA. MEDI-528 Clinical Trials Group. Safety profile and clinical activity of multiple subcutaneous doses of MEDI-528, a humanized anti-interleukin-9 monoclonal antibody, in two randomized phase 2a studies in subjects with asthma. BMC Pulm Med. 2011; 11:14.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Effects of CpG-oligodeoxynucleotides in Chronic Inflammation and Remodeling of Airway in a Murine Model of Bronchial Asthma
- TRPV1 Blocking Alleviates Airway Inflammation and Remodeling in a Chronic Asthma Murine Model
- Role of IL-23 and Th17 Cells in Airway Inflammation in Asthma
- The Effectiveness of Pimecrolimus in Airway Inflammation and Bronchial Hyperresponsiveness in Murine Asthma Model
- Effect of DHEA on airway hyperresponsiveness and inflammation in murine model of asthma
