Aberrant Epigenetic Modifications of LPHN2 Function as a Potential Cisplatin-Specific Biomarker for Human Gastrointestinal Cancer
- Affiliations
-
- 1Cancer Research Institute, Seoul National University, Seoul, Korea. kimty@snu.ac.kr
- 2Department of Molecular Medicine and Biopharmaceutical Sciences, Graduate School of Convergence Science and Technology, Seoul National University, Seoul, Korea.
- 3Department of Internal Medicine, Seoul National University Hospital, Seoul, Korea.
- KMID: 2454346
- DOI: http://doi.org/10.4143/crt.2015.153
Abstract
- PURPOSE
Epigenetic alterations of specific genes have recently been identified as diagnostic biomarkers for human cancers. However, there are currently no standardized epigenetic biomarkers for drug sensitivity in human gastrointestinal cancer. Therefore, the aim of this study is to identify a novel epigenetic biomarker in gastrointestinal cancer.
MATERIALS AND METHODS
Using bisulfite sequencing and pyrosequencing analysis, DNA methylation patterns of gastric, colon primary tissues and their cancer cells were analyzed, and histone modifications were analyzed using chromatin immunoprecipitation assay. In addition, cancer cells were exposed to cisplatin and treated with a DNA methyltransferase inhibitor.
RESULTS
We report that in human gastric and colon cancers, latrophilin 2 (LPHN2) is silenced by epigenetic modifications, including CpG island methylation and aberrant histone modifications. We also confirmed that LPHN2 was silenced by DNA hypermethylation in primary gastric and colon tumor tissues compared to their normal counterparts. Interestingly, we found that cancer cells with methylated LPHN2 showed higher sensitivity to cisplatin. Also, 5-aza- 2"²-deoxycytidine combined with cisplatin decreased the cytotoxicity of cisplatin in cancer cells with methylated LPHN2. In addition, LPHN2 knockdown in cancer cells with high LPHN2 expression sensitized these cells to the anti-proliferative effects of cisplatin.
CONCLUSION
In human gastrointestinal cancer, we found that LPHN2 is regulated by epigenetic modifications, and that cancer cells with lower LPHN2 expression show higher sensitivity to cisplatin. Therefore, the methylation status of LPHN2 is a potential novel epigenetic biomarker for cisplatin treatment in human gastric and colon cancers.
Keyword
MeSH Terms
Figure
-
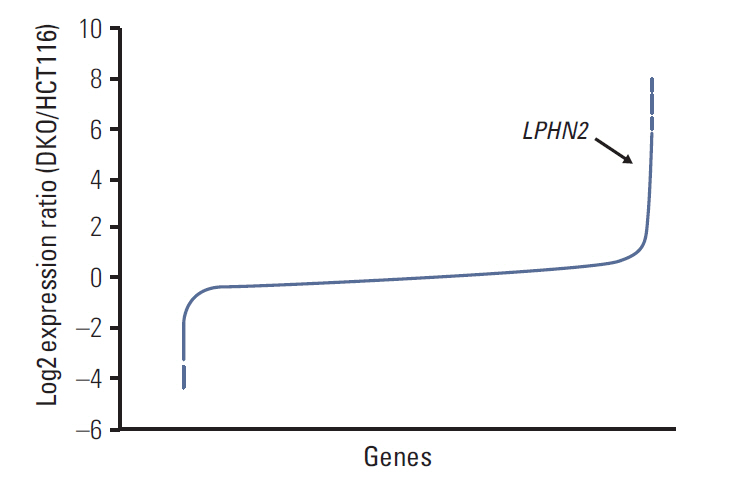
Fig. 1. Microarray analyses of mRNA expression in HCT116 and DKO cells. Microarray analyses were performed in HCT116 cells and DNMT1/DNMT3B double knockout derivative DKO cells. Microarray data results are shown as the log2 expression ratio of genes between HCT116 and DKO. The black arrow indicates LPHN2.

Fig. 2. DNA methylation and histone modifications of LPHN2 in human gastric and colon cancer cells. (A) Schematic representation of LPHN2. LPHN2 exons are indicated by gray boxes. Vertical bars represent each CpG site. The CpG island is indicated according to University of California Santa Cruz (UCSC) genome browser. (B) LPHN2 mRNA from 11 gastric cancer cell lines (left panel) and 15 colorectal cancer cell lines (right panel) was analyzed by reverse transcription polymerase chain reaction (RT-PCR). 18S ribosomal RNA served as an internal control. DW, distilled water. (C) Bisulfite genomic sequencing analyses of the LPHN2 CpG islands in SNU601, SNU484, HCT116, DKO, LOVO, and SW480 cells. Each row of squares denotes a single plasmid cloned and sequenced from polymerase chain reaction products amplified from bisulfitetreated gDNA. Open and filled squares represent unmethylated and methylated CpG sites, respectively. TSS, transcriptional start site. (D) Characterization of chromatin modification patterns around LPHN2. Chromatin immunoprecipitation (ChIP) assays were performed in SNU601, SNU484, HCT116, and DKO cells using antibodies specific for the active histone marker H3K4me3 and the inactive histone markers H3K27me2 and H3K9me3. Enrichment was measured using quantitative real-time RT-PCR. Error bars represent the standard error of the mean for triplicate chromatin preparations.
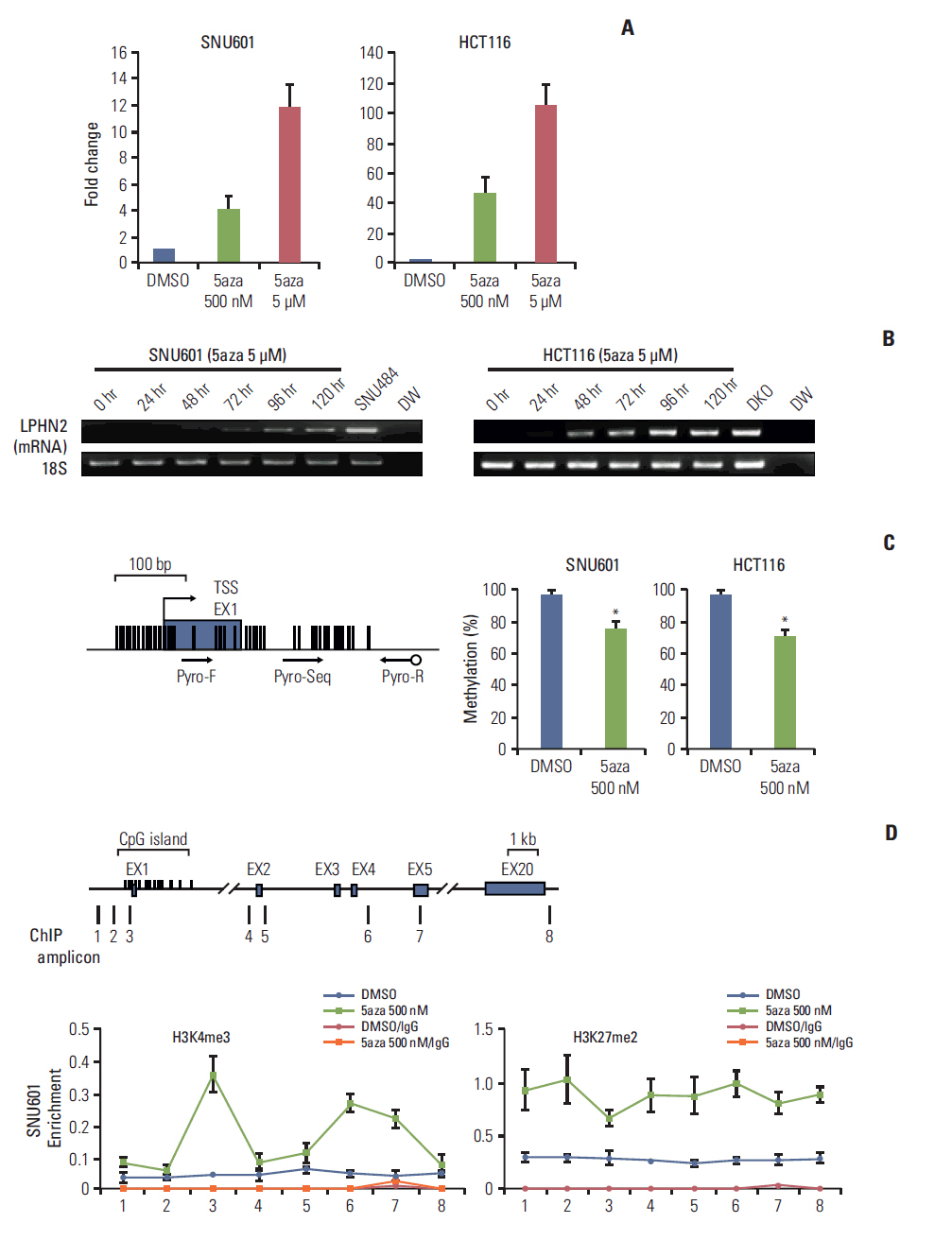
Fig. 3. Restoration of LPHN2 expression after treatment with 5-aza-2'-deoxycytidine (5-Aza-CdR). (A) SNU601 and HCT116 cells were treated with dimethyl sulfoxide (DMSO) or 5-Aza-CdR for 4 days. LPHN2 mRNA expression was measured using quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR). Each value was normalized to 18S ribosomal RNA. Error bars represent the standard deviation (SD) for three independent RNA preparations. (B) SNU601 and HCT116 cells were treated with 5 μM 5-Aza-CdR for 5 days. LPHN2 mRNA and 18S ribosomal RNA expression was analyzed by reverse transcription polymerase chain reaction at the indicated times. DW, distilled water. (C) After treatment with DMSO or 500 nM 5-Aza-CdR, the quantitative methylation status of LPHN2 in SNU601 and HCT116 cells was determined by pyrosequencing. The location of analyzed CpG sites corresponding to the pyrosequencing primer set is indicated by arrows. Error bars represent the SD for three independent preparations of bisulfite-treated gDNA. *p < 0.05 using Student’s t tests. (D) Dynamics of histone modifications after 5-Aza-CdR treatment in SNU601 cells. Chromatin was prepared from SNU601 cells treated with DMSO or 5-Aza-CdR (500 nM) for 4 days. Enrichment of the active histone mark (H3K4me3) and inactive histone mark (H3K27me2) was measured using qRT-PCR. Data are presented as the mean±standard error of the mean for three independent chromatin preparations. 5aza, 5-aza-2'-deoxycytidine.
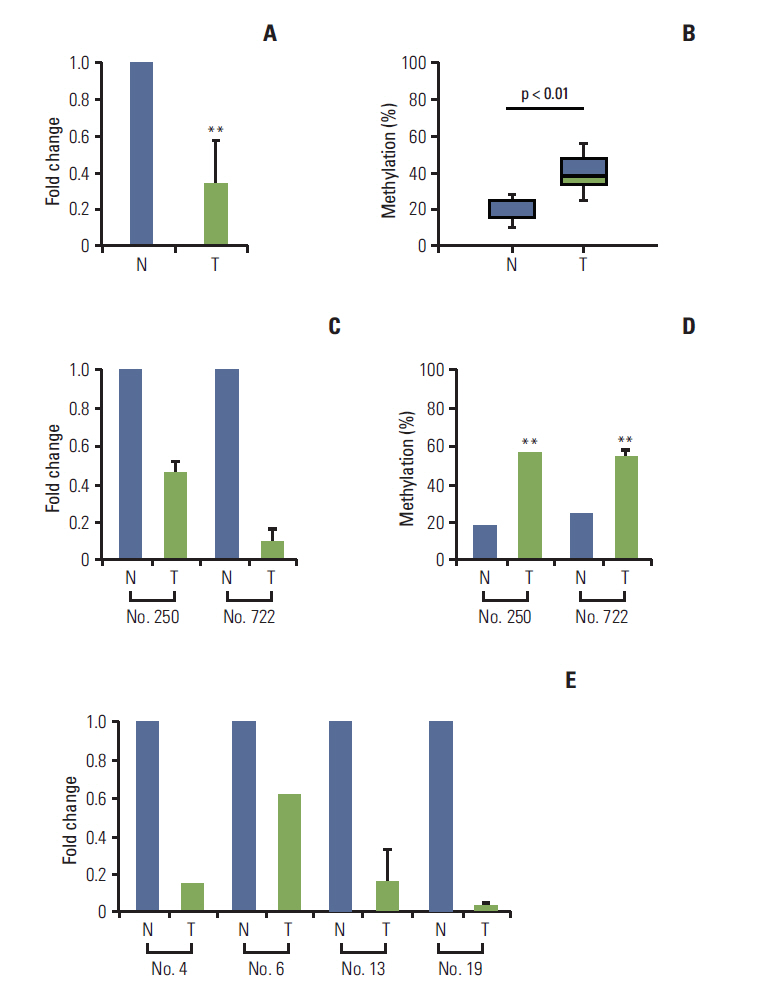
Fig. 4. Loss of LPHN2 expression in human gastric and colon primary tissues. mRNA expression in primary gastric tissues was analyzed using quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) and the methylation status of LPHN2 was confirmed via pyrosequencing analyses. Loss of LPHN2 mRNA expression was shown in 18 of 24 primary gastric tissues. (A) A summary of LPHN2 expression in 18 primary gastric tissues compared with normal tissues presented as fold change of relative mRNA expression. (B) Among 18 primary gastric tissues, nine of 18 showed hypermethylation of LPHN2 CpG islands compared to normal control tissues. The methylation ratio percentage is also presented as a box plot. (C) LPHN2 expression levels in primary gastric tissues were analyzed using qRT-PCR. LPHN2 expression was normalized to 18S ribosomal RNA and is shown as fold change relative to their normal tissue samples. Data are expressed as the mean±standard deivation. (D) Bisulfite-treated gDNAs from patient No. 250 and No. 722 were used to determine the methylation status of the LPHN2 promoter region using pyrosequencing analyses. (E) LPHN2 expression levels in primary colon tissues were analyzed by qRT-PCR and are shown as fold change relative to their normal tissue samples. N, normal tissue; T, tumor tissue. **p < 0.01 using Student’s t test.
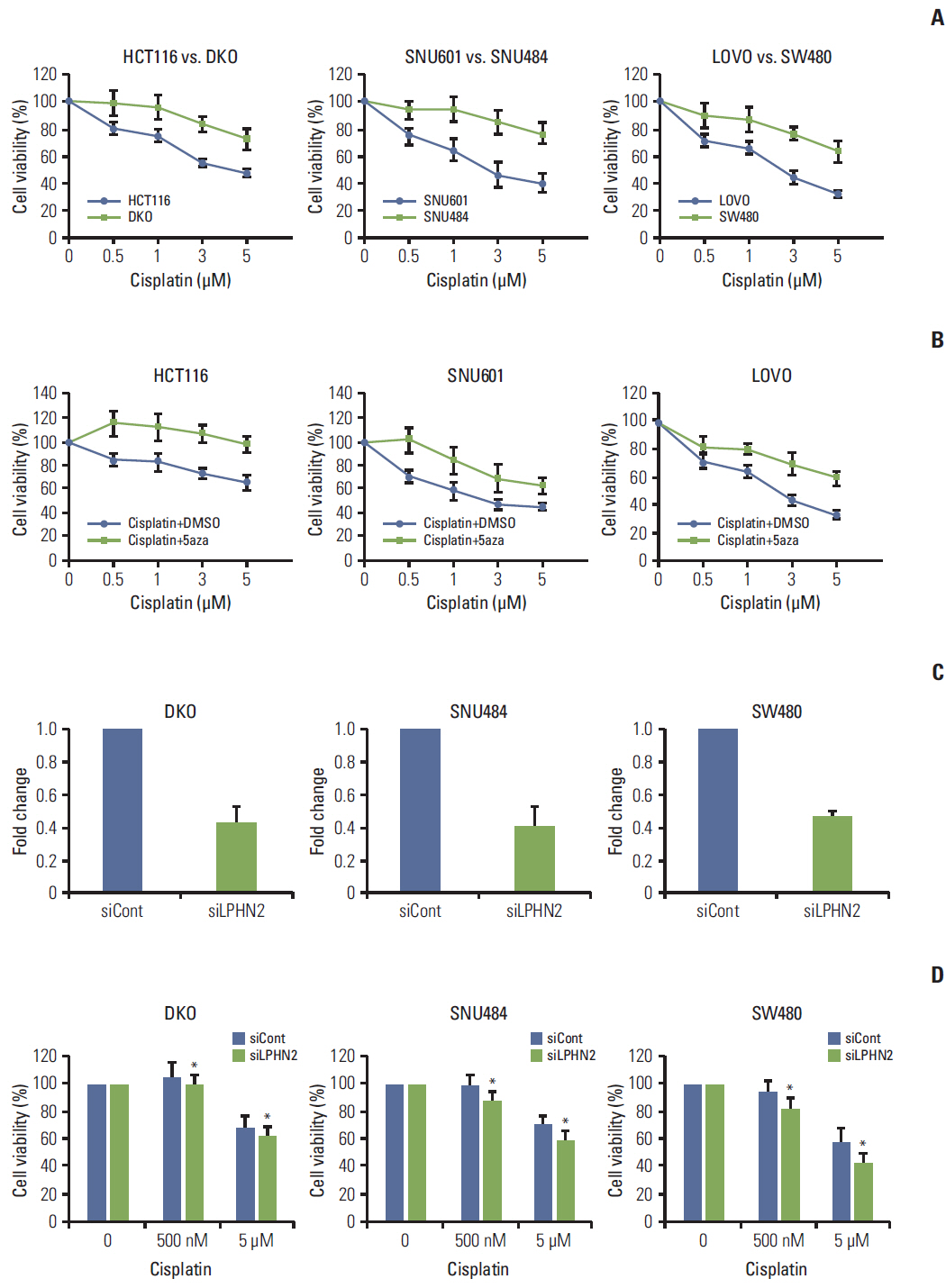
Fig. 5. Methylation status of LPHN2 determines the differential response to cisplatin. (A) Cells were treated with increasing concentrations of cisplatin (0.5, 1, 3, or 5 μM) for 72 hours. Cell viability was confirmed using MTT assays. Cell viability is expressed as a percentage relative to untreated cells. Data represent the mean±standard deviation (SD) of three independent experiments. (B) Cells were treated with dimethyl sulfoxide or 500 nM 5-aza-2'-deoxycytidine in combination with different doses of cisplatin (0.5, 1, 3, or 5 μM). The number of viable cells was measured after 72 hours. Data are expressed as the mean±SD (n=3). (C, D) Cells were transfected with control or LPHN2-specific siRNAs for 8 hours. Cells were plated in 96-well plates, followed by cisplatin treatment (0.5 μM or 5 μM) for 72 hours. LPHN2 expression (C) and cell viability (D) were measured using quantitative real-time reverse transcription polymerase chain reaction and MTT assays, respectively. Error bars represent the SD (n=3, *p < 0.05). 5aza, 5-aza-2'-deoxycytidine.
Reference
-
References
1. Jones PA, Baylin SB. The epigenomics of cancer. Cell. 2007; 128:683–92.
Article2. Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell. 2012; 150:12–27.
Article3. Hatziapostolou M, Iliopoulos D. Epigenetic aberrations during oncogenesis. Cell Mol Life Sci. 2011; 68:1681–702.
Article4. Das PM, Singal R. DNA methylation and cancer. J Clin Oncol. 2004; 22:4632–42.
Article5. Jung KW, Won YJ, Oh CM, Kong HJ, Cho H, Lee DH, et al. Prediction of cancer incidence and mortality in Korea, 2015. Cancer Res Treat. 2015; 47:142–8.
Article6. Hamilton JP, Meltzer SJ. A review of the genomics of gastric cancer. Clin Gastroenterol Hepatol. 2006; 4:416–25.
Article7. Nakajima T, Enomoto S, Ushijima T. DNA methylation: a marker for carcinogen exposure and cancer risk. Environ Health Prev Med. 2008; 13:8–15.
Article8. Naumov VA, Generozov EV, Zaharjevskaya NB, Matushkina DS, Larin AK, Chernyshov SV, et al. Genome-scale analysis of DNA methylation in colorectal cancer using Infinium Human-Methylation450 BeadChips. Epigenetics. 2013; 8:921–34.
Article9. Fang JY, Xiao SD. Alteration of DNA methylation in gastrointestinal carcinogenesis. J Gastroenterol Hepatol. 2001; 16:960–8.
Article10. Takeshima H, Wakabayashi M, Hattori N, Yamashita S, Ushijima T. Identification of coexistence of DNA methylation and H3K27me3 specifically in cancer cells as a promising target for epigenetic therapy. Carcinogenesis. 2015; 36:192–201.
Article11. Lelianova VG, Davletov BA, Sterling A, Rahman MA, Grishin EV, Totty NF, et al. Alpha-latrotoxin receptor, latrophilin, is a novel member of the secretin family of G protein-coupled receptors. J Biol Chem. 1997; 272:21504–8.12. Ku JL, Park JG. Biology of SNU cell lines. Cancer Res Treat. 2005; 37:1–19.
Article13. Rhee I, Bachman KE, Park BH, Jair KW, Yen RW, Schuebel KE, et al. DNMT1 and DNMT3b cooperate to silence genes in human cancer cells. Nature. 2002; 416:552–6.
Article14. Song SH, Kim A, Ragoczy T, Bender MA, Groudine M, Dean A. Multiple functions of Ldb1 required for beta-globin activation during erythroid differentiation. Blood. 2010; 116:2356–64.15. Yun J, Song SH, Park J, Kim HP, Yoon YK, Lee KH. Gene silencing of EREG mediated by DNA methylation and histone modification in human gastric cancers. Lab Invest. 2012; 92:1033–44.16. Song SH, Hou C, Dean A. A positive role for NLI/Ldb1 in long-range beta-globin locus control region function. Mol Cell. 2007; 28:810–22.17. Silva JP, Ushkaryov YA. The latrophilins, "split-personality" receptors. Adv Exp Med Biol. 2010; 706:59–75.
Article18. Rodriguez-Paredes M, Esteller M. Cancer epigenetics reaches mainstream oncology. Nat Med. 2011; 17:330–9.
Article19. Esteller M. Cancer epigenomics: DNA methylomes and histone-modification maps. Nat Rev Genet. 2007; 8:286–98.
Article20. Rodriguez J, Munoz M, Vives L, Frangou CG, Groudine M, Peinado MA. Bivalent domains enforce transcriptional memory of DNA methylated genes in cancer cells. Proc Natl Acad Sci USA. 2008; 105:19809–14.
Article21. Garritano S, Inga A, Gemignani F, Landi S. More targets, more pathways and more clues for mutant p53. Oncogenesis. 2013; 2:e54.
Article22. Zhang S, Liu Y, Liu Z, Zhang C, Cao H, Ye Y, et al. Transcriptome profiling of a multiple recurrent muscle-invasive urothelial carcinoma of the bladder by deep sequencing. PLoS One. 2014; 9:e91466.
Article23. Zheng CX, Gu ZH, Han B, Zhang RX, Pan CM, Xiang Y, et al. Whole-exome sequencing to identify novel somatic mutations in squamous cell lung cancers. Int J Oncol. 2013; 43:755–64.
Article24. Park KS, Kim HK, Lee JH, Choi YB, Park SY, Yang SH, et al. Transglutaminase 2 as a cisplatin resistance marker in non-small cell lung cancer. J Cancer Res Clin Oncol. 2010; 136:493–502.
Article25. Ivanova T, Zouridis H, Wu Y, Cheng LL, Tan IB, Gopalakrishnan V, et al. Integrated epigenomics identifies BMP4 as a modulator of cisplatin sensitivity in gastric cancer. Gut. 2013; 62:22–33.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Epigenetic Cross-Talk between DNA Methylation and Histone Modifications in Human Cancers
- Genetic and epigenetic alterations of colorectal cancer
- Epigenetic Modifications: Novel Therapeutic Approach for Thyroid Cancer
- Epigenetic Alterations and Loss of Imprinting in Colorectal Cancer
- Annexin A5 as a New Potential Biomarker for Cisplatin-Induced Toxicity in Human Kidney Epithelial Cells
