IKKγ Facilitates the Activation of NF-κB by Hepatitis C Virus Core Protein
- Affiliations
-
- 1Department of Biohealth Sciences, Changwon National University, Changwon, Korea. dwkim@changwon.ac.kr
- 2Department of Biology and Chemistry, Changwon National University Changwon, Korea.
- KMID: 2421304
- DOI: http://doi.org/10.4167/jbv.2018.48.3.93
Abstract
- Hepatitis C virus (HCV) is a major cause of chronic hepatitis, liver cirrhosis and hepatocellular carcinoma. HCV core protein has been shown to modulate various cellular signaling pathways including the nuclear factor κB (NF-κB) pathway which is associated with inflammation, cell proliferation and apoptosis. However, there have been conflicting reports about the effect of HCV core protein on NF-κB pathway, and the mechanism by which the core protein affects NF-κB activity remains nuclear. In this study, the functional interaction of HCV core protein and IκB kinase γ (IKKγ) was investigated using the expression plasmids of core and the components of IKK complex. The data revealed that HCV core protein activates NF-κB. Also, HCV core protein up-regulated the phosphorylation and degradation of IκBα. The activating effect of HCV core protein on NF-κB was synergistically elevated by IKKγ. It was noticed that the N-terminal IKKβ binding site, C-terminal leucine zipper, and zinc finger domains of IKKγ are not necessary for its synergistic effect. HCV core protein and IKKγ appeared to activate NF-κB by up-regulating the IKKβ activity resulting in the degradation of IκBα. As expected, HCV core protein induced the expression of NF-κB-targeted pro-inflammatory genes such as iNOS, IL-1β and IL-6 in the transcription level. These results suggest that HCV core protein induces NF-κB through the interaction with IKKγ and may play a critical role in the development of inflammation and related liver diseases.
Keyword
MeSH Terms
Figure
-
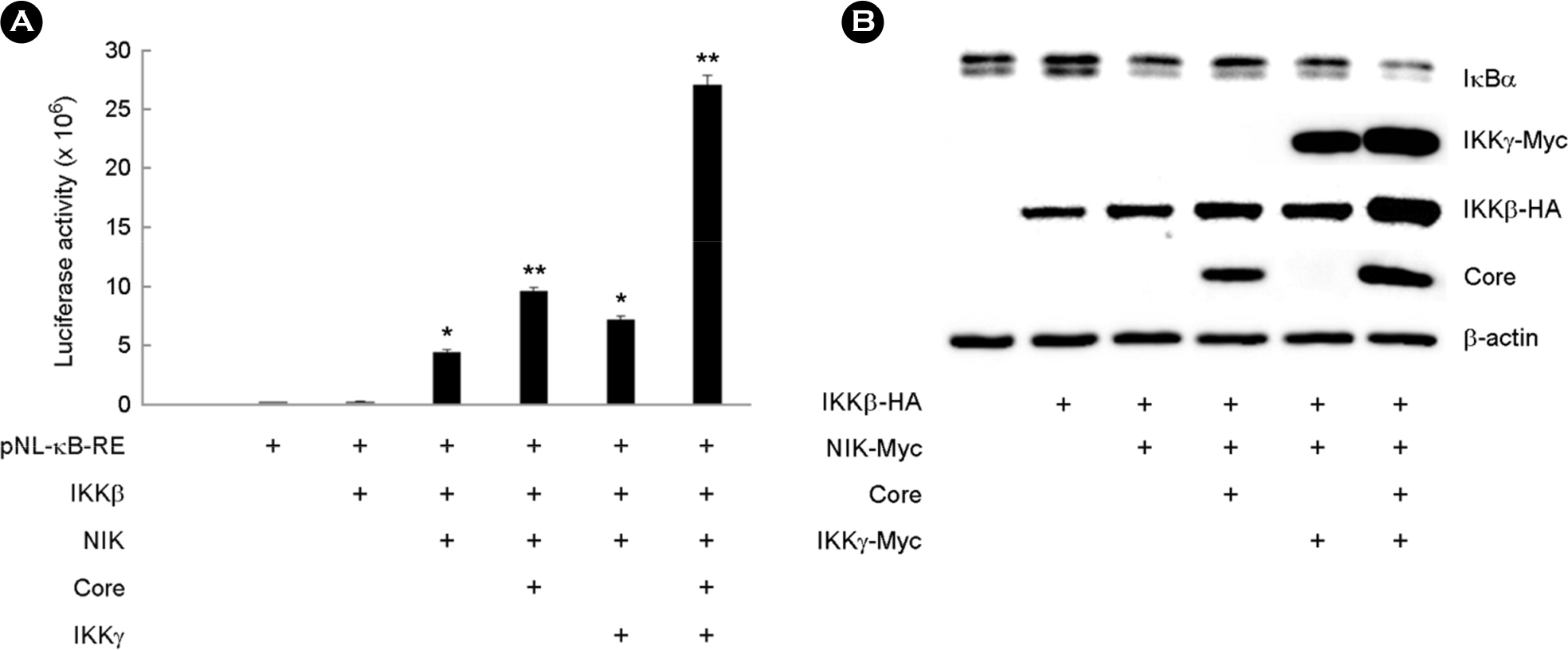
Figure 1. Activation of NF-κB by HCV core and IKKγ. (A) COS-7 cells were co-transfected with pNL-κB-RE reporter and each expression plasmid of IKKβ, NIK, HCV core, or IKKγ. Luciferase assays were performed 45 hr after transfection. (mean ± SD, n=3, ∗ p<0.05, ∗∗ p<0.001). (B) COS-7 cells were co-transfected with indicated expression plasmids. Cells were harvested at 45 hr post-transfection and the cytoplasmic extracts were analyzed by Western blotting.
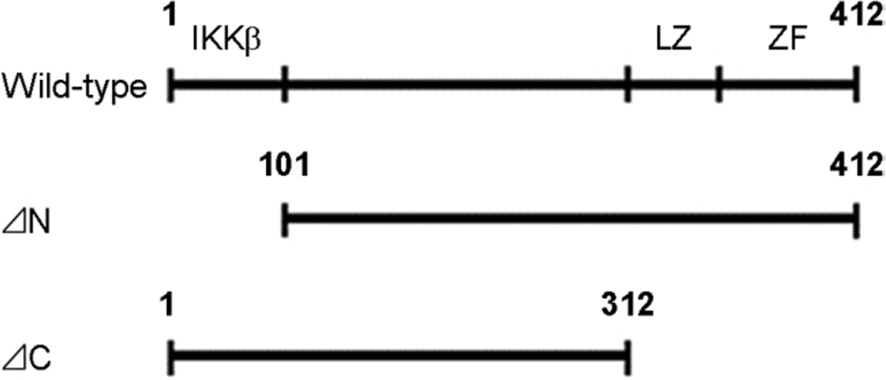
Figure 2. The structure of wild-type and N-terminal, C-terminal deletion mutants of mouse IKKγ. IKKβ binding domain at N-terminal and leucine zipper domain (LZ), zinc finger domain (ZF) at C-terminal are shown. The numbers indicate the number of amino acids.
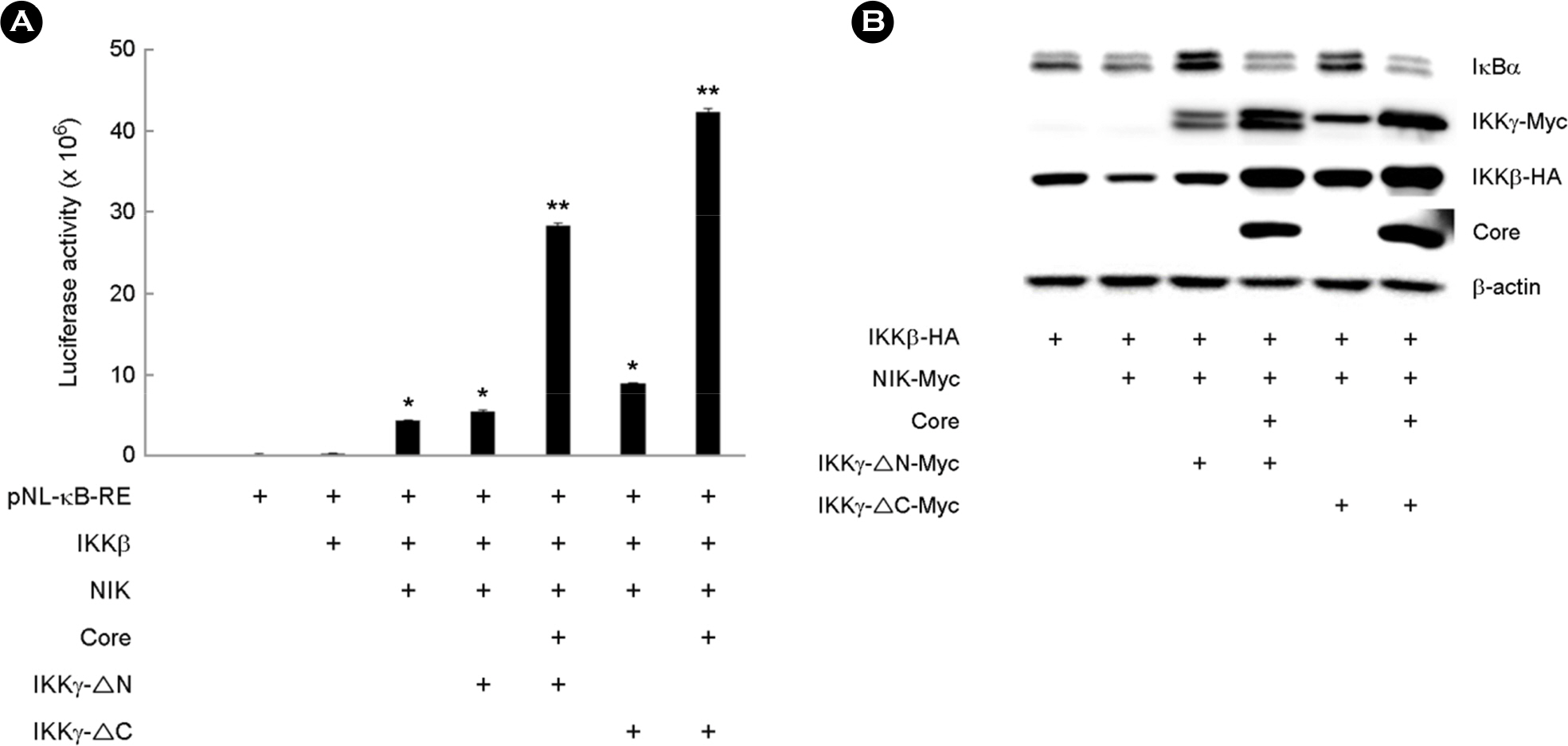
Figure 3. Activation of NF-κB by HCV core and IKKγ mutants. (A) COS-7 cells were co-transfected with pNL-κB-RE reporter and each expression plasmid of IKKβ, NIK, HCV core or IKKγ mutant. Luciferase assay was performed at 45 hr post-transfection. (mean ± SD, n=3, ∗ p<0.05, ∗∗ p<0.001). (B) COS-7 cells were co-transfected with expression plasmids as indicated. Cells were harvested at 45 hr post-transfection and the cytoplasmic extracts were analyzed by western blotting.
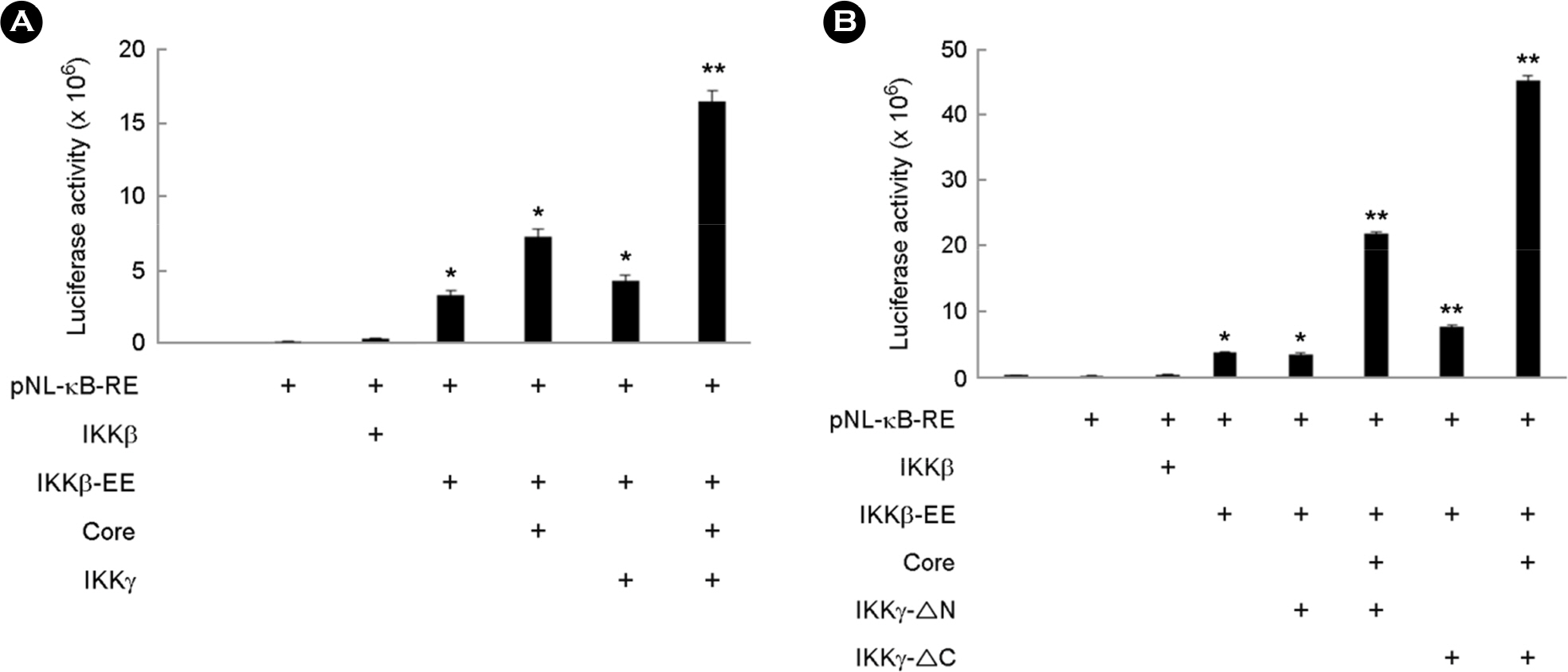
Figure 4. Effect of HCV core and IKKγ on the activation of NF-κB. COS-7 cells were co-transfected with pNL-κB-RE reporter and each expression plasmid of IKKβ, IKKβ-EE, HCV core, wild-type IKKγ, IKKγ-Δ N, or Δ C. Luciferase assay was performed at 45 hr post-transfection (mean ± SD, n=3, ∗ p<0.05, ∗∗ p<0.001).
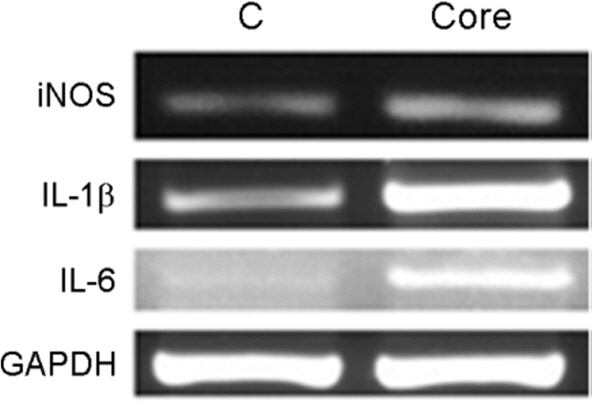
Figure 5. Effect of HCV core on the expression of inflammatory mediators. Raw 264.7 cells were transfected with empty vector (C) or HCV core-expression plasmid (core). Cells were harvested at 45 hr post-transfection and analyzed by RT-PCR. GAPDH was used as the internal control.
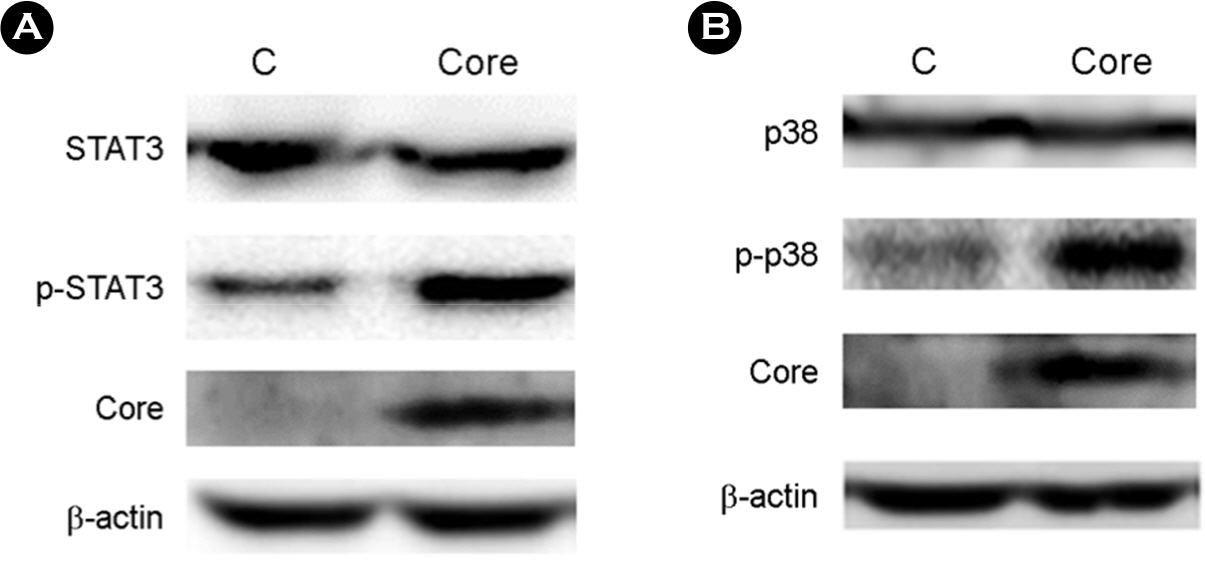
Figure 6. Effect of HCV core on the phosphorylation of signaling molecules in the proinflammatory pathway. Raw 264.7 cells were transfected with empty vector (C) or HCV core expression plasmid. The whole cell extracts were prepared at 45 hr post-transfection and analyzed by western blotting.
Reference
-
1). Tanaka K, Hirohata T, Koga S, Sugimachi K, Kanematsu T, Ohryohji F, et al. Hepatitis C and hepatitis B in the etiology of hepatocellular carcinoma in the Japanese population. Cancer Res. 1991; 51:2842–7.2). Grakoui A, Wychowski C, Lin C, Feinstone SM, Rice CM. Expression and identification of hepatitis C virus polyprotein cleavage products. J Virol. 1993; 67:1385–95.
Article3). Hijikata M, Kato N, Ootsuyama Y, Nakagawa M, Shimotohno K. Gene mapping of the putative structural region of the hepatitis C virus genome by in vitro processing analysis. Proc Natl Acad Sci USA. 1991; 88:5547–51.
Article4). Mizushima H, Hijikata M, Tanji Y, Kimura K, Shimotohno K. Analysis of N-terminal processing of hepatitis C virus nonstructural protein 2. J Virol. 1994; 68:2731–4.
Article5). Shrivastava A, Manna SK, Ray R, Aggarwal BB. Ectopic expression of hepatitis C virus core protein differentially regulates nuclear transcription factors. J Virol. 1998; 72:9722–8.
Article6). Yoshida H, Kato N, Shiratori Y, Otsuka M, Maeda S, Kato J, et al. Hepatitis C virus core protein activates nuclear factor κ B-dependent signaling through tumor necrosis factor receptor-associated factor. J Biol Chem. 2001; 276:16399–405.7). Marusawa H, Hijicata M, Chiba T, Shimotohno K. Hepatitis C virus core protein inhibits Fas- and tumor necrosis factor alpha-mediated apoptosis via NF-κB activation. J Virol. 1999; 73:4713–20.
Article8). Zhu N, Ware CF, Lai MM. Hepatitis C virus core protein enhances FADD-mediated apoptosis and suppresses TRADD signaling of tumor necrosis factor receptor. Virology. 2001; 283:178–87.
Article9). Park J, Kang W, Ryu SW, Kim WI, Chang DY, Lee DH, et al. Hepatitis C virus infection enhances TNFα-induced cell death via suppression of NF-κB. Hepatology. 2012; 56:831–40.
Article10). Ray RB, Meyer K, Ray R. Suppression of apoptotic cell death by hepatitis C virus core protein. Virology. 1996; 226:176–82.
Article11). Ghosh G, Wang VY, Huang DB, Fusco A. NF-κB regulation: lessons from structures. Immunol Rev. 2012; 246:36–58.
Article12). Hayden MS, Ghosh S. Shared principles in NF-κB signaling. Cell. 2008; 132:344–62.
Article13). Oeckinghaus A, Hayden MS, Ghosh S. Crosstalk in NF-κB signaling pathways. Nat Immunol. 2011; 12:695–708.
Article14). Ghosh S, Febin Prabhu Dass J. Non-canonical pathway network modelling and ubiquitination site prediction through homology modelling of NF-κB. Gene. 2016; 581:48–56.
Article15). Hinz M, Arslan SC, Scheidereit C. It takes two to tango: IκBs, the multifunctional partners of NF-κB. Immunol Rev. 2012; 246:59–76.
Article16). Morgan MJ, Liu ZG. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011; 21:103–15.
Article17). Mauro C, Pacifico F, Lavorgna A, Mellone S, Iannetti A, Acquaviva R, et al. ABIN-1 binds to NEMO/IKKγ and co-operates with A20 in inhibiting NF-κB. J Biol Chem. 2006; 281:18482–8.
Article18). Yamamoto Y, Kim DW, Kwak YT, Parajapati S, Verma U, Gaynor RB. IKKγ/NEMO facilitates the recruitment of the IκB proteins into the IκB kinase complex. J Biol Chem. 2001; 276:36327–36.
Article19). Kwon WJ, Kim SH, Park YO, Cho M, Kang CD, Lee G, et al. IKKγ inhibits activation of NF-κB by NIK. Mol cells. 2004; 18:200–6.20). Li XH, Fang X, Gaynor RB. Role of Ikkγ/NEMO in assembly of the ikappa B kinase complex. J Biol Chem. 2001; 276:4494–500.21). Mercurio F, Zhu H, Murray BW, Shevchenko A, Bennett BL, Li J, et al. Ikk-1 and Ikk-2: Cytokine-activated ikappa B kinase essential for NF-κB activation. Science. 1997:278:860–6.22). Tang H, Chen H, Jia Y, Liu X, Han Z, Wang A, et al. Effect of inhibitors of endocytosis and NF-κB signal pathway on folate-conjugated nanoparticle endocytosis by rat Kupffer cells. Int J Nanomedicine. 2017; 12:6937–47.23). He G, Karin M. NF-κB and STAT3 – key players in liver inflammation and cancer. Cell Res. 2011; 21:159–68.
Article24). Craig R, Larkin A, Mingo AM, Thuerauf DJ, Andrews C, McDonough PM, et al. p38 MAPK and NF-κB collaborate to induce interleukin-6 gene expression and release. J Biol Chem. 2000; 275:23814–24.
Article25). Bouffard P, Hayashi PH, Acevedo R, Levy N, Zeldis JB. Hepatitis C virus is detected in a monocyte/macrophage subpopulation of peripheral blood mononuclear cells of infected patients. J Infect Dis. 1992; 166:1276–80.
Article26). Heydtmann M, Adams DH. Chemokines in the immunopathogenesis of hepatitis C infection. Hepatology. 2009; 49:676–88.
Article27). Saito K, Meyer K, Warner R, Basu A, Ray RB, Ray R. Hepatitis C virus core protein inhibits tumor necrosis factor alpha-mediated apoptosis by a protective effect involving cellular FLICE inhibitory protein. J virol. 2006; 80:4372–9.
Article28). McLauchlan J. Properties of the hepatitis C virus core protein: a structural protein that modulates cellular processes. J viral hepat. 2000; 7:2–14.
Article29). Bühler S, Bartenschlager R. Promotion of hepatocellular carcinoma by hepatitis C virus. Dig Dis. 2012; 30:445–52.
Article30). Hayden MS, Ghosh S. NF-κB in immunology. Cell Res. 2011; 21:223–44.31). Xing Y, Wang X, Jameson SC, Hogquist KA. Late stage of T cell maturation in the thymus involve NF-κB and tonic type I interferon signaling. Nat Immunol. 2016; 17:565–73.32). Colotta F, Allavena P, Sica A, Garlanda C, Mantovani A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis. 2009; 30:1073–81.
Article33). Bours V, Bentires-Alj M, Hellin AC, Viatour P, Robe P, Delhalle S, et al. Nuclear factor-kappa B, cancer, and apoptosis. Biochem Pharmacol. 2000; 60:1085–9.34). Wu JT, Kral JG. The NF-kappaB/IkappaB signaling system: A molecular target in breast cancer therapy. J Surg Res. 2005; 123:158–69.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Immunohistochemical Analysis for Basal Activation of NF-κB in Acral Lentiginous Melanoma
- Controls of Nuclear Factor-Kappa B Signaling Activity by 5’-AMP-Activated Protein Kinase Activation With Examples in Human Bladder Cancer Cells
- Induction of Interleukin-8 Expression in Synovial Cell by Hepatitis C Virus Core Protein
- Prevention of Viral Hepatitis and Vaccination
- New biomarkers of hepatitis B virus (HBV) infection: HBV RNA and HBV core-related antigen, new kids on the block?
