Myometrial relaxation of mice via expression of two pore domain acid sensitive K⺠(TASK-2) channels
- Affiliations
-
- 1Department of Obstetrics and Gynecology, Chungbuk National University College of Medicine, Cheongju 28644, Korea. iwji@chungbuk.ac.kr
- 2Department Physiology, Chungbuk National University College of Medicine, Cheongju 28644, Korea. physiokyc@chungbuk.ac.kr
- 3Department of Pharmacology, Chungbuk National University College of Medicine, Cheongju 28644, Korea.
- 4Department of Urology, College of Medicine, Chung-Ang University, Seoul 06974, Korea.
- 5Department of Physiology, College of Medicine, Chung-Ang University, Seoul 06974, Korea.
- 6VHS Medical Center, Seoul 05368, Korea.
- 7Department of Physiology, Nagoya City University Medical School, Nagoya 467-8601, Japan.
- 8Department of Obstetrics and Gynecology, Cheongju St. Mary's Hospital, Cheongju 28323, Korea.
- 9Department of Preventive Medicine, Chungbuk National University College of Medicine, Cheongju 28644, Korea.
- 10Department of Anesthesiology and Pain Medicine, Chungbuk National University College of Medicine, Cheongju 28644, Korea.
- 11Department of Physiology, Shanghai Jiaotong University, School of Medicine, Shanghai, 200240, P.R. China.
- 12Department of Medical Education, Chungbuk National University College of Medicine, Cheongju 28644, Korea.
- KMID: 2350512
- DOI: http://doi.org/10.4196/kjpp.2016.20.5.547
Abstract
- Myometrial relaxation of mouse via expression of two-pore domain acid sensitive (TASK) channels was studied. In our previous report, we suggested that two-pore domain acid-sensing K⺠channels (TASK-2) might be one of the candidates for the regulation of uterine circular smooth muscles in mice. In this study, we tried to show the mechanisms of relaxation via TASK-2 channels in marine myometrium. Isometric contraction measurements and patch clamp technique were used to verify TASK conductance in murine myometrium. Western blot and immunehistochemical study under confocal microscopy were used to investigate molecular identity of TASK channel. In this study, we showed that TEA and 4-AP insensitive non-inactivating outward K⺠current (NIOK) may be responsible for the quiescence of murine pregnant longitudinal myometrium. The characteristics of NIOK coincided with two-pore domain acid-sensing K⺠channels (TASK-2). NIOK in the presence of K⺠channel blockers was inhibited further by TASK inhibitors such as quinidine, bupivacaine, lidocaine, and extracellular acidosis. Furthermore, oxytocin and estrogen inhibited NIOK in pregnant myometrium. When compared to non-pregnant myometrium, pregnant myometrium showed stronger inhibition of NIOK by quinidine and increased immunohistochemical expression of TASK-2. Finally, TASK-2 inhibitors induced strong myometrial contraction even in the presence of L-methionine, a known inhibitor of stretch-activated channels in the longitudinal myometrium of mouse. Activation of TASK-2 channels seems to play an essential role for relaxing uterus during pregnancy and it might be one of the alternatives for preventing preterm delivery.
Keyword
MeSH Terms
Figure
-
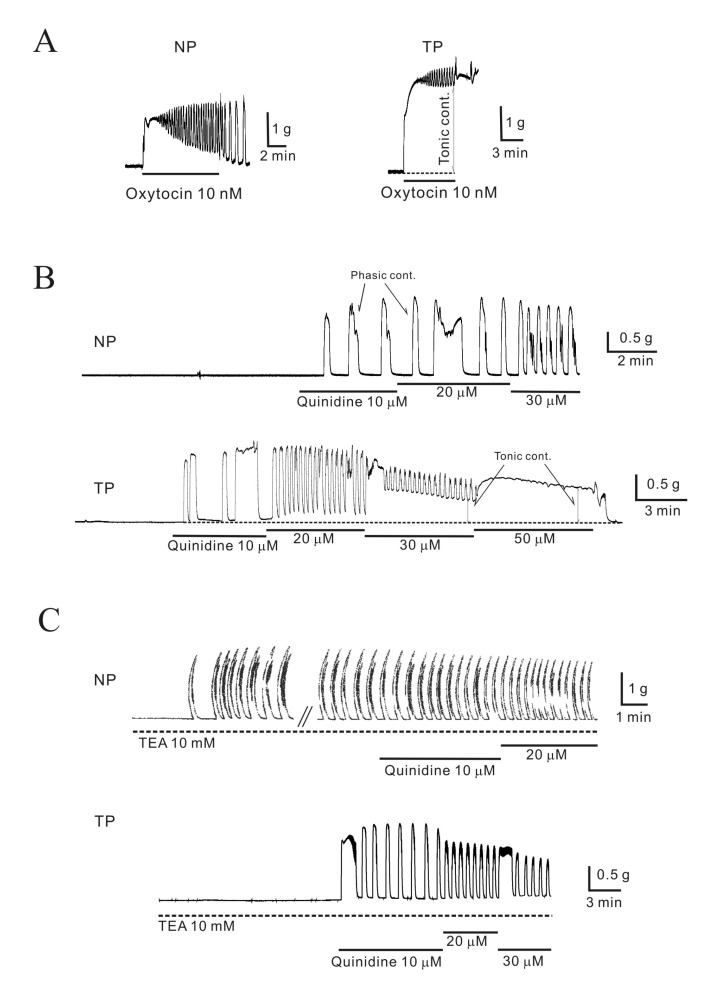
Fig. 1 Effect of quinidine on the isometric contraction of uterine longitudinal muscle in mice.(A) Oxytocin (OXT) produced initial and tonic contractions; however, tonic contractions were not sustained in non-pregnant myometrium (NP, non-pregnant; TP, term-pregnant). (B) Quinidine (10, 20, and 30 µM) produced concentration-dependent phasic contractions. Tonic contractions were also produced at high concentrations of quinidine in pregnant myometrium. (C) Quinidine was applied to myometrium in the presence of TEA (10 mM). TEA (10 mM) produced strong phasic contractions in non-pregnant myometrium (1.2±0.15 g) but not in pregnant myometrium. Quinidine (10~30 µM), even in the presence of TEA, increased contraction frequency in non-pregnant myometrium and produced stronger contractions in pregnant myometrium.
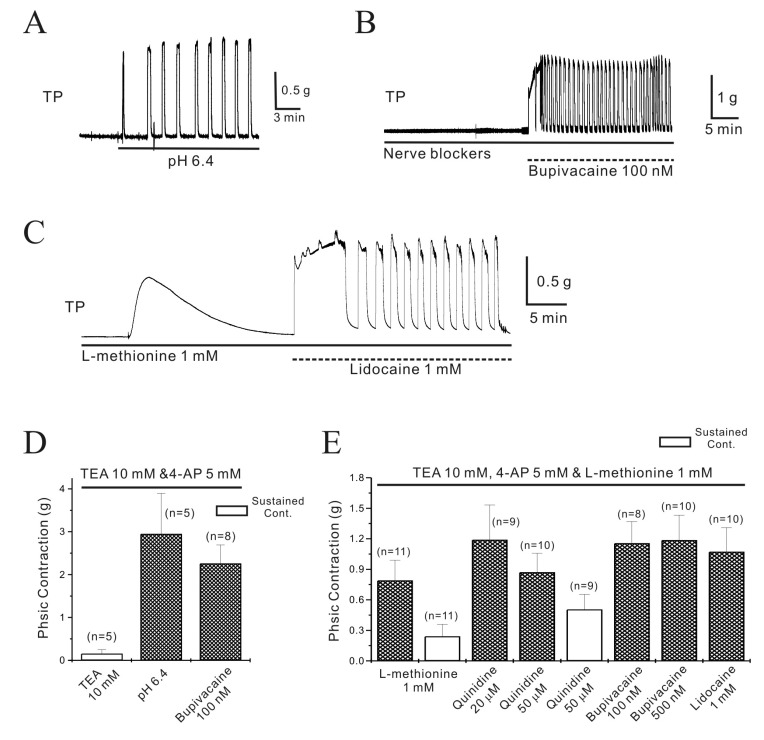
Fig. 2 Regulation of pregnant mouse myometrial contractility by TASK-2 channel inhibitors in the presence of TEA and 4-AP.The TASK-2 inhibitors produced contractions in pregnant myometrium in the presence of TEA and 4-AP. (A) Extracellular acidosiso (pHo=6.4) produced contractions in pregnant myometrium. (B) Bupivacaine (100 nM) also produced robust contractions even in the presence of nerve blockers. (C) L-methionine (1 mM), which inhibits stretch-dependent K2P channels (TREK-1), produced contractions that spontaneously decayed to near baseline values within 10~15 min. However, lidocaine produced robust contraction in the presence of L-methionine. (D~E) Data are summarized.
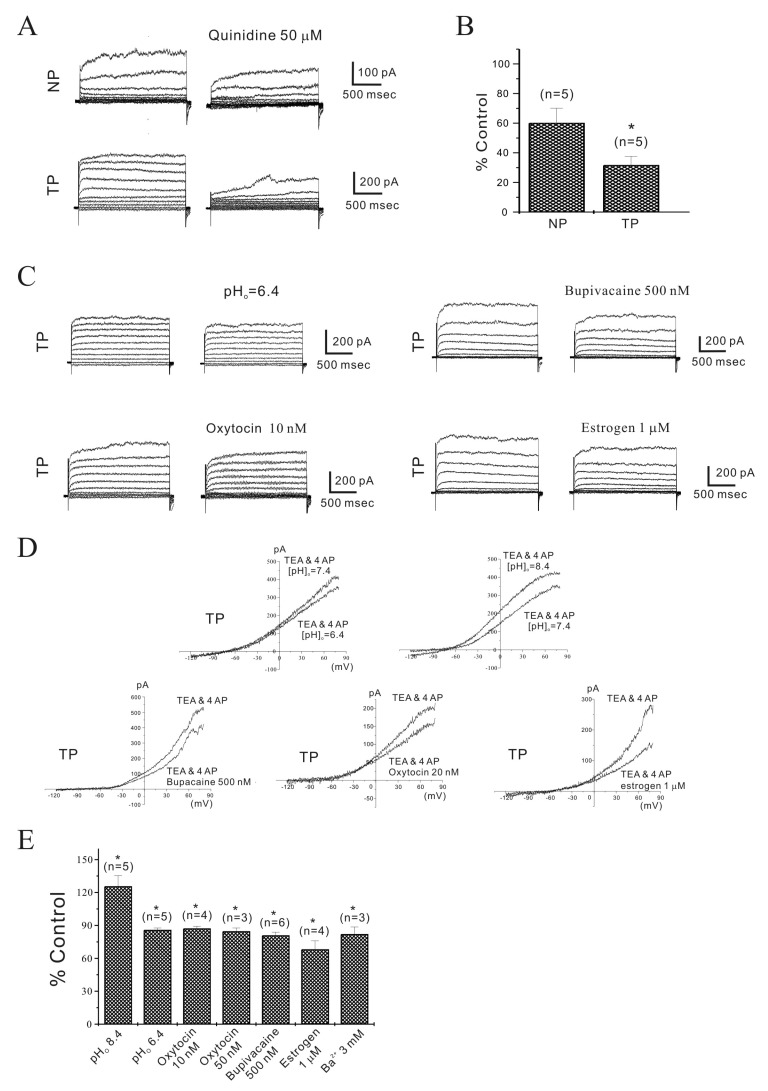
Fig. 3 Regulation of TEA and 4-AP insensitive non-inactivating outward current (NIOK) in single mouse myometrial cells.(A, B) In the presence of TEA (10 mM) and 4-AP (5 mM), NIOK were observed under whole-cell voltage clamp. Quinidine (50 µM) inhibited NIOK currents more significantly in pregnant cells. (C) NIOK was inhibited by pHo=6.4, bupivacaine, oxytocin (OXT), and estrogen in pregnant myometrial cells. (D) Representative I/V relationships are shown. (E) Data are summarized. NIOK was inhibited to 126, 86, 87, 85, 81, 68 and 78% of the control by pHo=8.4, pHo=6.4, OXT (10 and 50 nM), bupivacaine (500 nM), estrogen (1 µM), and Ba2+ (3 mM), respectively (n= 5, 5, 4, 3, 6, 4 and 3, respectively; *p<0.05). Such as TASK-2 is very promising for managing labor at term and developing target medicines. As shown in Fig. 3C~3E, OXT (10 and 50 nM) inhibited the NIOK current to 87±1.94% and 85±2.93% in pregnant cells in a reversible manner, respectively (p<0.05). Estrogen (1 µM) also inhibited the NIOK current to 68±7.65% (n=4; p<0.05).
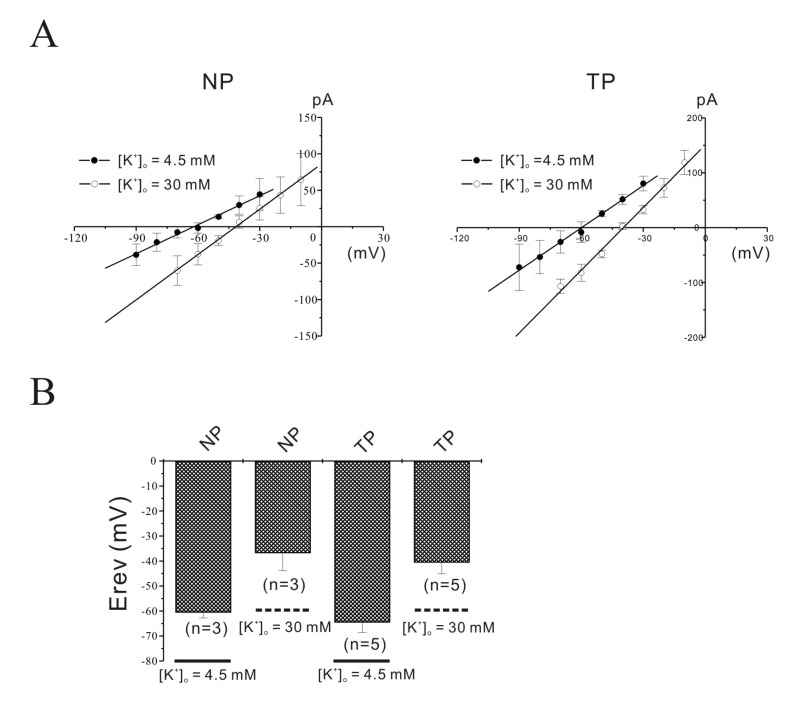
Fig. 4 Identification of NIOK by reversal potentials (Erev).The reversal potentials (Erev) of NIOK were evaluated by changing the external K+ concentration from 4.5 to 30 mM. Erev shifted to new expected values at 30 mM of external K+. Instantaneous tail currents in the NIOK of non-pregnant and pregnant myometrial cells were reversed at –37 mV (n=3) and –41 mV (n=5) by 30 mM K+.
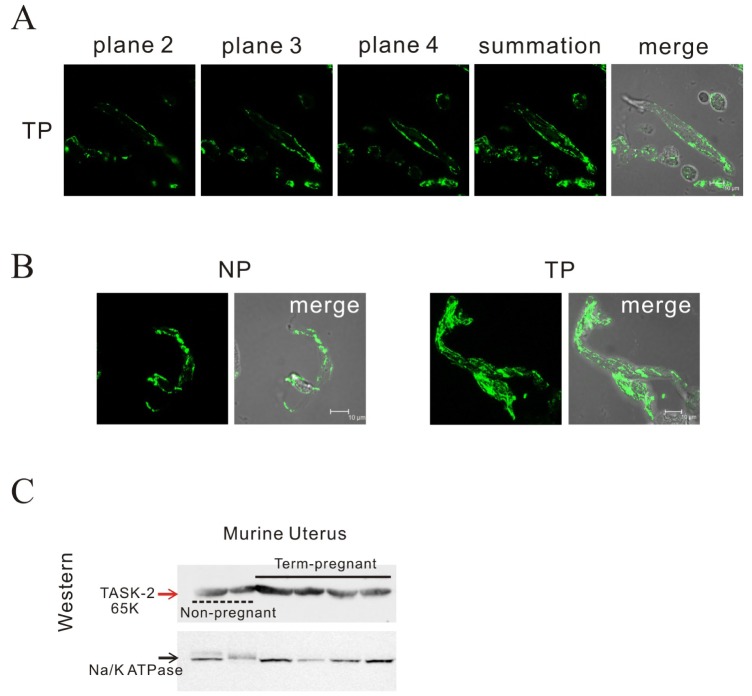
Fig. 5 Identification of TASK-2 by immunohistochemistry and Western blot in mouse myometrium.(A, B) Laser scanning confocal micrograph shows TASK-2 immunoreactivity on the membrane of a single pregnant cell. Confocal images at 2 µm intervals showed localization of the TASK-2 channel on the cell membrane of pregnant myometrial cells. Highly increased immunoreactivity for the TASK-2 antibody in single pregnant myometrial cells ws observed compared to that in non-pregnant cells. (C) TASK-2 was increased in pregnancy compared to the control under Western blot (595 vs 100, p<0.05; n=3 and 4).
Reference
-
1. Parkington HC, Coleman HA. Excitability in uterine smooth muscle. Front Horm Res. 2001; 27:179–200. PMID: 11450426.
Article2. Sanborn BM. Ion channels and the control of myometrial electrical activity. Semin Perinatol. 1995; 19:31–34. PMID: 7538700.
Article3. Garfield RE, Sims S, Daniel EE. Gap junctions: their presence and necessity in myometrium during parturition. Science. 1977; 198:958–960. PMID: 929182.
Article4. Wray S, Noble K. Sex hormones and excitation-contraction coupling in the uterus: the effects of oestrous and hormones. J Neuroendocrinol. 2008; 20:451–461. PMID: 18266942.
Article5. Cerri V, Tarantini M, Zuliani G, Schena V, Redaelli C, Nicolini U. Intravenous glucose infusion in labor does not affect maternal and fetal acid-base balance. J Matern Fetal Med. 2000; 9:204–208. PMID: 11048829.
Article6. Sjostedt S. Acid-base balance of arterial blood during pregnancy, at delivery, and in the puerperium. Am J Obstet Gynecol. 1962; 84:775–779. PMID: 13913556.7. Harrison N, Larcombe-McDouall JB, Earley L, Wray S. An in vivo study of the effects of ischaemia on uterine contraction, intracellular pH and metabolites in the rat. J Physiol. 1994; 476:349–354. PMID: 8046648.
Article8. Brinkman CR. Circulation in the pregnant uterus. In : Carsten ME, Miller JD, editors. Uterine function: molecular and cellular aspects. New York: Plenum Press;1990. p. 519–537.10. Tulchinsky D, Hobel CJ, Yeager E, Marshall JR. Plasma estrone, estradiol, estriol, progesterone, and 17-hydroxyprogesterone in human pregnancy. I. Normal pregnancy. Am J Obstet Gynecol. 1972; 112:1095–1100. PMID: 5025870.11. Walsh SW, Stanczyk FZ, Novy MJ. Daily hormonal changes in the maternal, fetal, and amniotic fluid compartments before parturition in a primate species. J Clin Endocrinol Metab. 1984; 58:629–639. PMID: 6230368.
Article12. Hong SH, Kyeong KS, Kim CH, Kim YC, Choi W, Yoo RY, Kim HS, Park YJ, Ji IW, Jeong EH, Kim HS, Xu WX, Lee SJ. Regulation of myometrial contraction by ATP-sensitive potassium (KATP) channel via activation of SUR2B and Kir 6.2 in mouse. J Vet Med Sci. 2016; 78:1153–1159. PMID: 27086859.13. Park JK, Kim YC, Sim JH, Choi MY, Choi W, Hwang KK, Cho MC, Kim KW, Lim SW, Lee SJ. Regulation of membrane excitability by intracellular pH (pHi) changers through Ca2+-activated K+ current (BK channel) in single smooth muscle cells from rabbit basilar artery. Pflugers Arch. 2007; 454:307–319. PMID: 17285302.14. Smith RC, McClure MC, Smith MA, Abel PW, Bradley ME. The role of voltage-gated potassium channels in the regulation of mouse uterine contractility. Reprod Biol Endocrinol. 2007; 5:41–52. PMID: 17980032.
Article15. Wray S, Burdyga T, Noble D, Noble K, Borysova L, Arrowsmith S. Progress in understanding electro-mechanical signalling in the myometrium. Acta Physiol (Oxf). 2015; 213:417–431. PMID: 25439280.
Article16. Anwer K, Oberti C, Perez GJ, Perez-Reyes N, McDougall JK, Monga M, Sanborn BM, Stefani E, Toro L. Calcium-activated K+ channels as modulators of human myometrial contractile activity. Am J Physiol. 1993; 265:C976–C985. PMID: 8238323.17. Brayden JE, Nelson MT. Regulation of arterial tone by activation of calcium-dependent potassium channels. Science. 1992; 256:532–535. PMID: 1373909.
Article18. Bai X, Bugg GJ, Greenwood SL, Glazier JD, Sibley CP, Baker PN, Taggart MJ, Fyfe GK. Expression of TASK and TREK, two-pore domain K+ channels, in human myometrium. Reproduction. 2005; 129:525–530. PMID: 15798028.19. Ketchum KA, Joiner WJ, Sellers AJ, Kaczmarek LK, Goldstein SA. A new family of outwardly rectifying potassium channel proteins with two pore domains in tandem. Nature. 1995; 376:690–695. PMID: 7651518.
Article20. O'Connell AD, Morton MJ, Hunter M. Two-pore domain K+ channels-molecular sensors. Biochim Biophys Acta. 2002; 1566:152–161. PMID: 12421546.21. Hong SH, Sung R, Kim YC, Suzuki H, Choi W, Park YJ, Ji IW, Kim CH, Myung SC, Lee MY, Kang TM, You RY, Lee KJ, Lim SW, Yun HY, Song YJ, Xu WX, Kim HS, Lee SJ. Mechanism of relaxation via TASK-2 channels in uterine circular muscle of mouse. Korean J Physiol Pharmacol. 2013; 17:359–365. PMID: 23946696.
Article22. Duprat F, Lesage F, Fink M, Reyes R, Heurteaux C, Lazdunski M. TASK, a human background K+ channel to sense external pH variations near physiological pH. EMBO J. 1997; 16:5464–5471. PMID: 9312005.23. Kim YC, Suzuki H, Xu WX, Hashitani H, Choi W, Yun HY, Park SM, Youn SJ, Lee SJ, Lee SJ. Voltage-dependent Ca2+ current identified in freshly isolated interstitial clls of cajal (ICC) of Guineapig stomach. Korean J Physiol Pharmacol. 2008; 12:323–330. PMID: 19967074.24. Reyes R, Duprat F, Lesage F, Fink M, Salinas M, Farman N, Lazdunski M. Cloning and expression of a novel pH-sensitive two pore domain K+ channel from human kidney. J Biol Chem. 1998; 273:30863–30869. PMID: 9812978.25. Buxton IL, Singer CA, Tichenor JN. Expression of stretch-activated two-pore potassium channels in human myometrium in pregnancy and labor. PLoS One. 2010; 5:e12372. PMID: 20811500.
Article26. Monaghan K, Baker SA, Dwyer L, Hatton WC, Sik Park K, Sanders KM, Koh SD. The stretch-dependent potassium channel TREK-1 and its function in murine myometrium. J Physiol. 2011; 589:1221–1233. PMID: 21224218.
Article27. Miller P, Kemp PJ, Lewis A, Chapman CG, Meadows HJ, Peers C. Acute hypoxia occludes hTREK-1 modulation: re-evaluation of the potential role of tandem P domain K+ channels in central neuroprotection. J Physiol. 2003; 548:31–37. PMID: 12611912.28. Kang D, Mariash E, Kim D. Functional expression of TRESK-2, a new member of the tandem-pore K+ channel family. J Biol Chem. 2004; 279:28063–28070. PMID: 15123670.29. Che T, Sun H, Li J, Yu X, Zhu D, Xue B, Liu K, Zhang M, Kunze W, Liu C. Oxytocin hyperpolarizes cultured duodenum myenteric intrinsic primary afferent neurons by opening BK(Ca) channels through IP3 pathway. J Neurochem. 2012; 121:516–525. PMID: 22356163.30. Soriano S, Ripoll C, Fuentes E, Gonzalez A, Alonso-Magdalena P, Ropero AB, Quesada I, Nadal A. Regulation of K(ATP) channel by 17β-estradiol in pancreatic β-cells. Steroids. 2011; 76:856–860. PMID: 21470558.
Article31. Druzin M, Malinina E, Grimsholm O, Johansson S. Mechanism of estradiol-induced block of voltage-gated K+ currents in rat medial preoptic neurons. PLoS One. 2011; 6:e20213. PMID: 21625454.32. Ma XY, Yu JM, Zhang SZ, Liu XY, Wu BH, Wei XL, Yan JQ, Sun HL, Yan HT, Zheng JQ. External Ba2+ block of the two-pore domain potassium channel TREK-1 defines conformational transition in its selectivity filter. J Biol Chem. 2011; 286:39813–39822. PMID: 21965685.33. Naderali EK, Wray S. Modulation of force induced by pH in the guinea-pig uterus examined at two stages of the oestrous cycle. J Reprod Fertil. 1999; 117:153–157. PMID: 10645256.
Article34. Taggart MJ, Sheader EA, Walker SD, Naderali EK, Moore S, Wray S. External alkalinization decreases intracellular Ca++ and spontaneous contractions in pregnant rat myometrium. Am J Obstet Gynecol. 1997; 177:959–963. PMID: 9369852.
Article35. Rooth G, Sjostedt S. The placental transfer of gases and fixed acids. Arch Dis Child. 1962; 37:366–370. PMID: 14493734.
Article36. Parratt JR, Taggart MJ, Wray S. Functional effects of intracellular pH alteration in the human uterus: simultaneous measurements of pH and force. J Reprod Fertil. 1995; 105:71–75. PMID: 7490717.
Article37. Taggart M, Wray S. Simultaneous measurement of intracellular pH and contraction in uterine smooth muscle. Pflugers Arch. 1993; 423:527–529. PMID: 8351202.
Article38. Soloff MS, Fernstrom MA, Periyasamy S, Soloff S, Baldwin S, Wieder M. Regulation of oxytocin receptor concentration in rat uterine explants by estrogen and progesterone. Can J Biochem Cell Biol. 1983; 61:625–630. PMID: 6313158.
Article39. Mesiano S, Welsh TN. Steroid hormone control of myometrial contractility and parturition. Semin Cell Dev Biol. 2007; 18:321–333. PMID: 17613262.
Article40. Khan RN, Matharoo-Ball B, Arulkumaran S, Ashford ML. Potassium channels in the human myometrium. Exp Physiol. 2001; 86:255–264. PMID: 11429642.
Article41. Khan RN, Smith SK, Morrison JJ, Ashford ML. Ca2+ dependence and pharmacology of large-conductance K+ channels in nonlabor and labor human uterine myocytes. Am J Physiol. 1997; 273:C1721–C1731. PMID: 9374660.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- DAMGO modulates two-pore domain K⺠channels in the substantia gelatinosa neurons of rat spinal cord
- Volatile General Anesthetics and Tandem Pore Domain K+ Channel
- Two-pore Domain Potassium Channels in Astrocytes
- A Deafness Associated Protein TMEM43 Interacts with KCNK3 (TASK-1) Two-pore Domain K+ (K2P) Channel in the Cochlea
- Identification of ATP-sensitive K+ Channel in an Uterine Leiomyoma Cell
