Antiangiogenic Activity of Acer tegmentosum Maxim Water Extract in Vitro and in Vivo
- Affiliations
-
- 1Division of Biological Science and Technology and Yonsei-Fraunhofer Medical Device Lab., College of Science and Technology, Yonsei University, Wonju, Korea.
- 2Division of Endocrinology and Metabolism, Department of Internal Medicine, Inha University School of Medicine, Incheon, Korea. namms@inha.ac.kr
- 3Qingdao Endocrine & Diabetes Hospital, Qingdao, China.
- 4Department of Biomedical Engineering, Institute of Medical Engineering and Yonsei-Fraunhofer Medical Device Laboratory, Yonsei University, Wonju, Korea.
- 5Department of Biomedical Sciences, College of Medicine, Inha University, Incheon, Korea.
- 6Department of Obstetrics & Gynecology, Inha University College of Medicine, Incheon, Korea.
- 7Department of Clinical Pharmacology, Inha University School of Medicine, Incheon, Korea.
- KMID: 2164486
- DOI: http://doi.org/10.3346/jkms.2015.30.7.979
Abstract
- Angiogenesis, the formation of new blood vessels, is critical for tumor growth and metastasis. Notably, tumors themselves can lead to angiogenesis by inducing vascular endothelial growth factor (VEGF), which is one of the most potent angiogenic factors. Inhibition of angiogenesis is currently perceived as one of the most promising strategies for the blockage of tumor growth. In this study, we investigated the effects of Acer tegmentosum maxim water extract (ATME) on angiogenesis and its underlying signal mechanism. We studied the antiangiogenic activity of ATME by using human umbilical vein endothelial cells (HUVECs). ATME strongly inhibited VEGF-induced endothelial cell proliferation, migration, invasion, and tube formation, as well as vessel sprouting in a rat aortic ring sprouting assay. Moreover, we found that the p44/42 mitogen activated protein (MAP) kinase signaling pathway is involved in the inhibition of angiogenesis by ATME. Moreover, when we performed the in vivo matrigel plug assay, VEGF-induced angiogenesis was potently reduced when compared to that for the control group. Taken together, these results suggest that ATME exhibits potent antiangiogenic activity in vivo and in vitro and that these effects are regulated by the extracellular regulated kinase (ERK) pathway.
Keyword
MeSH Terms
-
Acer/*metabolism
Angiogenesis Inhibitors/*pharmacology
Animals
Cell Line, Tumor
Cell Movement/drug effects
Cell Proliferation/drug effects
Cell Survival
Extracellular Signal-Regulated MAP Kinases/*metabolism
Hep G2 Cells
Human Umbilical Vein Endothelial Cells/*drug effects
Humans
MAP Kinase Signaling System/drug effects
Mice
Mice, Inbred C57BL
Mitogen-Activated Protein Kinase 1/metabolism
Neoplasm Invasiveness/pathology
Neovascularization, Pathologic/*drug therapy/prevention & control
Nitric Oxide Synthase Type III/metabolism
Phosphorylation/drug effects
Plant Extracts/pharmacology
Rats
Rats, Sprague-Dawley
Transcription Factors/metabolism
Vascular Endothelial Growth Factor A/antagonists & inhibitors/metabolism
Angiogenesis Inhibitors
Extracellular Signal-Regulated MAP Kinases
Mitogen-Activated Protein Kinase 1
Nitric Oxide Synthase Type III
Plant Extracts
Transcription Factors
Vascular Endothelial Growth Factor A
Figure
-

Fig. 1 Effects of ATME on the cytotoxicity of HUVECs, MIAPaCa-2, HepG2, and CT-26 cells. HUVECs (A), MIAPaCa-2 (B), HepG2 (C), and CT-26 (D) cells were incubated with various concentrations (0.05, 0.1, 0.2, 0.5, and 1 mg/mL) of ATME. After 48 hr, cytotoxicity was quantified by an MTT assay. Each bar represents the average±SE of three independent experiments.
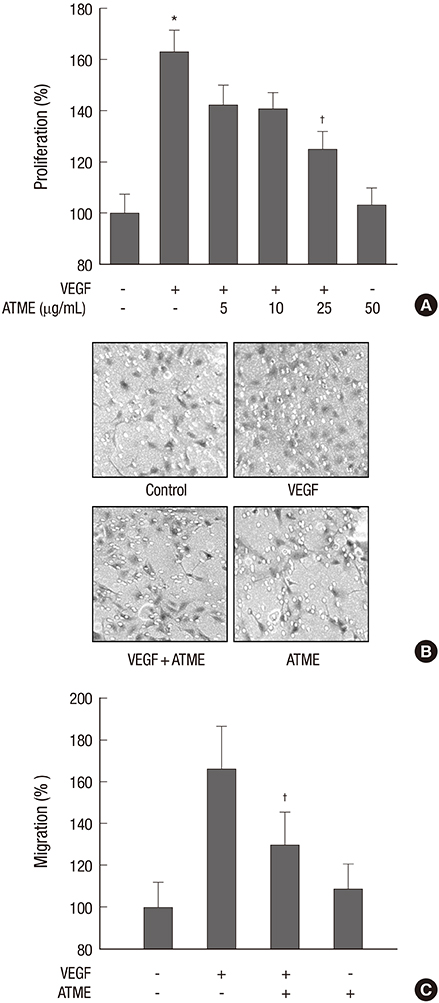
Fig. 2 ATME inhibits the VEGF-induced proliferation and migration of HUVECs. (A) HUVECs were pretreated for 30 min with various concentrations (5, 10, and 25 µg/mL) of ATME before exposure to VEGF (20 ng/mL). After 48 hr, the number of cells was determined by an MTT assay. (B, C) HUVECs were treated with VEGF (20 ng/mL) in the presence or absence of ATME (25 µg/mL). Chemotactic migration after incubation in Transwell plates for 4 hr. (B) Representative migrated cells were photographed. (C) Cells that migrated to the bottom of the filter were counted using optical microscopy. The in vitro angiogenesis assay was performed as described in the Materials and Methods. Data are expressed as the means+SE (n = 3). *P < 0.01 vs. control and †P < 0.01 vs. VEGF alone.

Fig. 3 ATME inhibits VEGF-induced invasion and tube formation of endothelial cells. (A) Effect of ATME on HUVEC invasion by using the Transwell culture plate. HUVECs were treated for 16 hr with VEGF (20 ng/mL) and various concentration (5, 10, 25, and 50 µg/mL) of ATME. VEGF treatment alone served as a positive control. (B, C) HUVECs were preincubated for 30 min with 25 µg/mL ATME and plated on Matrigel-coated plates at a density of 2×105 cells per well. They were incubated in the presence or absence of 20 ng/mL VEGF, and microphotographs were obtained after 20 hr (40×). (B) Representative endothelial tubes are shown. (C) The area covered by the tube network was measured using the Image-Pro Plus software. The experiments were repeated three times, and values are the means+SE values of triplicate determinations. *P < 0.01 vs. VEGF alone.
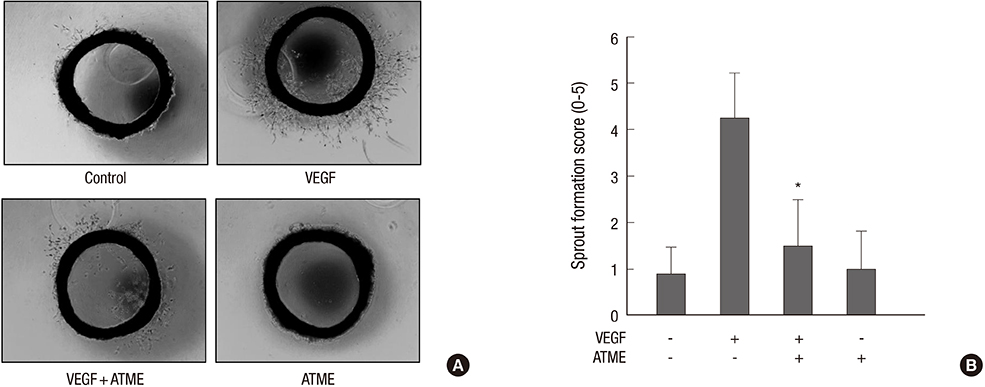
Fig. 4 ATME inhibits VEGF-induced vessel sprouting ex vivo. Aortae in Matrigel were exposed to VEGF (20 ng/mL) in the absence or presence of ATME (25 µg/mL) and stained with Diff-Quick on day 7. (A) Representative aortic rings were photographed. (B) ATME blocks VEGF-induced vessel sprouting. The assay was scored from 0 (least positive) to 5 (most positive), and the data are mean±SE values (n = 6). *P < 0.01 vs. VEGF alone.
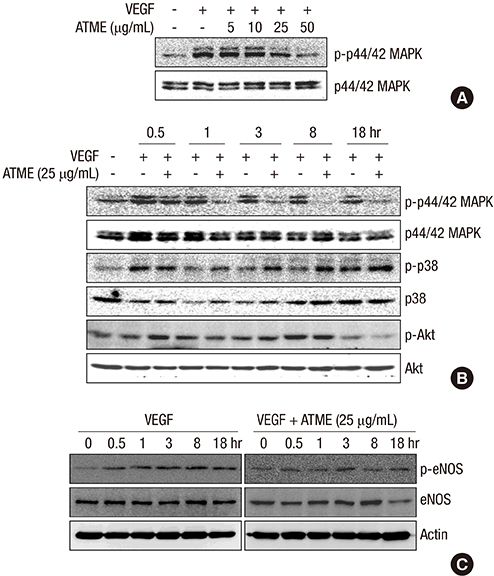
Fig. 5 ATME inhibits VEGF-induced phosphorylation of p44/42 MAP kinase and eNOS but not phosphorylation of p38 and Akt. (A) HUVECs were pretreated for 30 min with various concentrations (5, 10, 25, and 50 µg/mL) of ATME and treated with VEGF (20 ng/mL) for 30 min. (B and C) HUVECs were incubated with 25 µg/mL ATME for 30 min and stimulated with VEGF (20 ng/mL) for the indicated times. At the indicated time points, cells were harvested and the levels of phosphorylated and total p44/42 MAP kinase, p38, Akt, and eNOS were determined by western blot analysis. Actin was used as a loading control.
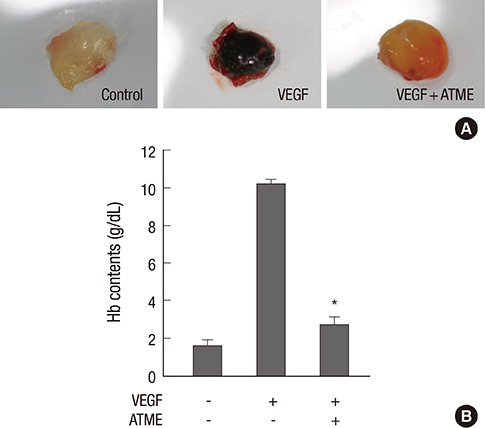
Fig. 6 Angiogenesis was reduced by ATME in the in vivo mouse Matrigel-plus assay. The experimental procedures are described under Materials and Methods. (A) The Matrigel without ATME did not show any inhibition of VEGF-induced migration or invasion of endothelial cells. However, with Matrigel containing ATME (25 µg), many blood vessels disappeared in the gel. (B) Quantitation of active vasculature inside the Matrigel by measurement of hemoglobin content. Each value represents the mean value for at least four animals, and similar results were obtained in three different experiments; bars, + SE. *P < 0.01 vs. VEGF alone.
Reference
-
1. Folkman J, Cotran R. Relation of vascular proliferation to tumor growth. Int Rev Exp Pathol. 1976; 16:207–248.2. Folkman J, Shing Y. Angiogenesis. J Biol Chem. 1992; 267:10931–10934.3. Reinke JM, Sorg H. Wound repair and regeneration. Eur Surg Res. 2012; 49:35–43.4. Veikkola T, Alitalo K. VEGFs, receptors and angiogenesis. Semin Cancer Biol. 1999; 9:211–220.5. McMahon G. VEGF receptor signaling in tumor angiogenesis. Oncologist. 2000; 5:3–10.6. Senger DR, Perruzzi CA, Feder J, Dvorak HF. A highly conserved vascular permeability factor secreted by a variety of human and rodent tumor cell lines. Cancer Res. 1986; 46:5629–5632.7. Plate KH, Breier G, Weich HA, Risau W. Vascular endothelial growth factor is a potential tumour angiogenesis factor in human gliomas in vivo. Nature. 1992; 359:845–848.8. Kim KJ, Li B, Winer J, Armanini M, Gillett N, Phillips HS, Ferrara N. Inhibition of vascular endothelial growth factor-induced angiogenesis suppresses tumour growth in vivo. Nature. 1993; 362:841–844.9. Manley PW, Bold G, Brüggen J, Fendrich G, Furet P, Mestan J, Schnell C, Stolz B, Meyer T, Meyhack B, et al. Advances in the structural biology, design and clinical development of VEGF-R kinase inhibitors for the treatment of angiogenesis. Biochim Biophys Acta. 2004; 1697:17–27.10. Sagar SM, Yance D, Wong RK. Natural health products that inhibit angiogenesis: a potential source for investigational new agents to treat cancer-Part 1. Curr Oncol. 2006; 13:14–26.11. Huang S, Yang N, Liu Y, Hu L, Zhao J, Gao J, Li Y, Li C, Zhang X, Huang T. Grape seed proanthocyanidins inhibit angiogenesis via the downregulation of both vascular endothelial growth factor and angiopoietin signaling. Nutr Res. 2012; 32:530–536.12. Lee SJ, Park K, Ha SD, Kim WJ, Moon SK. Gleditsia sinensis thorn extract inhibits human colon cancer cells: the role of ERK1/2, G2/M-phase cell cycle arrest and p53 expression. Phytother Res. 2010; 24:1870–1876.13. Yi JM, Park JS, Oh SM, Lee J, Kim J, Oh DS, Bang OS, Kim NS. Ethanol extract of Gleditsia sinensis thorn suppresses angiogenesis in vitro and in vivo. BMC Complement Altern Med. 2012; 12:243.14. Ahn DK. Illustrated book of Korean medicinal herbs. Seoul: Kyohak Publishing Co;1998.15. Morikawa T, Tao J, Toguchida I, Matsuda H, Yoshikawa M. Structures of new cyclic diarylheptanoids and inhibitors of nitric oxide production from Japanese folk medicine Acer nikoense. J Nat Prod. 2003; 66:86–91.16. Yonezawa T, Lee JW, Akazawa H, Inagaki M, Cha BY, Nagai K, Yagasaki K, Akihisa T, Woo JT. Osteogenic activity of diphenyl ether-type cyclic diarylheptanoids derived from Acer nikoense. Bioorg Med Chem Lett. 2011; 21:3248–3251.17. Barbehenn RV, Jones CP, Karonen M, Salminen JP. Tannin composition affects the oxidative activities of tree leaves. J Chem Ecol. 2006; 32:2235–2251.18. Park KM, Yang MC, Lee KH, Kim KR, Choi SU, Lee KR. Cytotoxic phenolic constituents of Acer tegmentosum maxim. Arch Pharm Res. 2006; 29:1086–1090.19. Jaffe EA, Nachman RL, Becker CG, Minick CR. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J Clin Invest. 1973; 52:2745–2756.20. Lee OH, Kim YM, Lee YM, Moon EJ, Lee DJ, Kim JH, Kim KW, Kwon YG. Sphingosine 1-phosphate induces angiogenesis: its angiogenic action and signaling mechanism in human umbilical vein endothelial cells. Biochem Biophys Res Commun. 1999; 264:743–750.21. Nicosia RF, Ottinetti A. Modulation of microvascular growth and morphogenesis by reconstituted basement membrane gel in three-dimensional cultures of rat aorta: a comparative study of angiogenesis in matrigel, collagen, fibrin, and plasma clot. In Vitro Cell Dev Biol. 1990; 26:119–128.22. Lal BK, Varma S, Pappas PJ, Hobson RW 2nd, Durán WN. VEGF increases permeability of the endothelial cell monolayer by activation of PKB/akt, endothelial nitric-oxide synthase, and MAP kinase pathways. Microvasc Res. 2001; 62:252–262.23. Ali N, Yoshizumi M, Fujita Y, Izawa Y, Kanematsu Y, Ishizawa K, Tsuchiya K, Yano S, Sone S, Tamaki T. A novel Src kinase inhibitor, M475271, inhibits VEGF-induced human umbilical vein endothelial cell proliferation and migration. J Pharmacol Sci. 2005; 98:130–141.24. Lee SJ, Namkoong S, Kim YM, Kim CK, Lee H, Ha KS, Chung HT, Kwon YG, Kim YM. Fractalkine stimulates angiogenesis by activating the Raf-1/MEK/ERK- and PI3K/Akt/eNOS-dependent signal pathways. Am J Physiol Heart Circ Physiol. 2006; 291:H2836–H2846.25. Birney YA, Sweeney CH, Cappadona CR, Sitzmann JV, Cummins PM, Redmond EM, Cahill PA. Pulse pressure-induced transmural fluid flux increases bovine aortic smooth muscle cell apoptosis in a mitogen activated protein kinase dependent manner. J Vasc Res. 2004; 41:364–374.26. Junttila MR, Li SP, Westermarck J. Phosphatase-mediated crosstalk between MAPK signaling pathways in the regulation of cell survival. FASEB J. 2008; 22:954–965.27. Huang C, Jacobson K, Schaller MD. MAP kinases and cell migration. J Cell Sci. 2004; 117:4619–4628.28. Finlay D, Healy V, Furlong F, O'Connell FC, Keon NK, Martin F. MAP kinase pathway signalling is essential for extracellular matrix determined mammary epithelial cell survival. Cell Death Differ. 2000; 7:302–313.29. Beckner ME. Factors promoting tumor angiogenesis. Cancer Invest. 1999; 17:594–623.30. Rousseau S, Houle F, Landry J, Huot J. p38 MAP kinase activation by vascular endothelial growth factor mediates actin reorganization and cell migration in human endothelial cells. Oncogene. 1997; 15:2169–2177.31. Chrzanowska-Wodnicka M, Kraus AE, Gale D, White GC 2nd, Vansluys J. Defective angiogenesis, endothelial migration, proliferation, and MAPK signaling in Rap1b-deficient mice. Blood. 2008; 111:2647–2656.32. Ziche M, Morbidelli L, Masini E, Amerini S, Granger HJ, Maggi CA, Geppetti P, Ledda F. Nitric oxide mediates angiogenesis in vivo and endothelial cell growth and migration in vitro promoted by substance P. J Clin Invest. 1994; 94:2036–2044.33. Babaei S, Stewart DJ. Overexpression of endothelial NO synthase induces angiogenesis in a co-culture model. Cardiovasc Res. 2002; 55:190–200.34. Morbidelli L, Chang CH, Douglas JG, Granger HJ, Ledda F, Ziche M. Nitric oxide mediates mitogenic effect of VEGF on coronary venular endothelium. Am J Physiol. 1996; 270:H411–H415.35. Ziche M, Morbidelli L, Choudhuri R, Zhang HT, Donnini S, Granger HJ, Bicknell R. Nitric oxide synthase lies downstream from vascular endothelial growth factor-induced but not basic fibroblast growth factor-induced angiogenesis. J Clin Invest. 1997; 99:2625–2634.36. Papapetropoulos A, García-Cardeña G, Madri JA, Sessa WC. Nitric oxide production contributes to the angiogenic properties of vascular endothelial growth factor in human endothelial cells. J Clin Invest. 1997; 100:3131–3139.37. Hood JD, Meininger CJ, Ziche M, Granger HJ. VEGF upregulates ecNOS message, protein, and NO production in human endothelial cells. Am J Physiol. 1998; 274:H1054–H1058.38. Kim YM, Namkoong S, Yun YG, Hong HD, Lee YC, Ha KS, Lee H, Kwon HJ, Kwon YG, Kim YM. Water extract of Korean red ginseng stimulates angiogenesis by activating the PI3K/Akt-dependent ERK1/2 and eNOS pathways in human umbilical vein endothelial cells. Biol Pharm Bull. 2007; 30:1674–1679.39. Lee PC, Salyapongse AN, Bragdon GA, Shears LL 2nd, Watkins SC, Edington HD, Billiar TR. Impaired wound healing and angiogenesis in eNOS-deficient mice. Am J Physiol. 1999; 277:H1600–H1608.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Analysis of α-Glucosidase Inhibitory Constituents from Acer tegmentosum Using LC-QTOF MS/MS And Molecular Networking
- HPLC and GC-MS Analysis of Phenolic Substances in Acer tegmentosum
- Acer tegmentosum Maxim Prevents Bone Loss by Inhibiting Osteoclastogenesis and Promoting Osteoblast Mineralization in Ovariectomized Mice
- Acer mono Extract Inhibits Invasive Activities and G1/S Transition of HT1080 Fibrosarcoma Cells
- 6-O-Galloylsalidroside, an Active Ingredient from Acer tegmentosum, Ameliorates Alcoholic Steatosis and Liver Injury in a Mouse Model of Chronic Ethanol Consumption
