Stanniocalcin-1 protects bovine intestinal epithelial cells from oxidative stress-induced damage
- Affiliations
-
- 1College of Veterinary Medicine, Huazhong Agricultural University, Wuhan 430070, China. vetwu@qq.com
- 2Hubei Key Laboratory of Animal Embryo and Molecular Breeding, Hubei Academy of Agricultural Science, Wuhan 430064, China.
- 3Biotechnology Institute of Animal and Veterinary Science, Sichan Animal Science Academy, Chengdu, 610066, China.
- KMID: 2070233
- DOI: http://doi.org/10.4142/jvs.2014.15.4.475
Abstract
- Chronic enteritis can produce an excess of reactive oxygen species resulting in cellular damage. Stanniocalcin-1(STC-1) reportedly possesses anti-oxidative activity, the aim of this study was to define more clearly the direct contribution of STC-1 to anti-oxidative stress in cattle. In this study, primary intestinal epithelial cells (IECs) were exposed to hydrogen peroxide (H2O2) for different time intervals to mimic chronic enteritis-induced cellular damage. Prior to treatment with 200 microM H2O2, the cells were transfected with a recombinant plasmid for 48 h to over-express STC-1. Acridine orange/ethidium bromide (AO/EB) double staining and trypan blue exclusion assays were then performed to measure cell viability and apoptosis of the cells, respectively. The expression of STC-1 and apoptosis-related proteins in the cells was monitored by real-time PCR and Western blotting. The results indicated that both STC-1 mRNA and protein expression levels positively correlated with the duration of H2O2 treatment. H2O2 damaged the bovine IECs in a time-dependent manner, and this effect was attenuated by STC-1 over-expression. Furthermore, over-expression of STC-1 up-regulated Bcl-2 protein expression and slightly down-regulated caspase-3 production in the damaged cells. Findings from this study suggested that STC-1 plays a protective role in intestinal cells through an antioxidant mechanism.
Keyword
MeSH Terms
-
Animals
Animals, Newborn
Blotting, Western/veterinary
Caspase 3/*genetics/metabolism
Cattle
Cattle Diseases/etiology/*genetics/metabolism
Duodenum/metabolism
Enteritis/etiology/genetics/metabolism/*veterinary
Epithelial Cells/metabolism
*Gene Expression Regulation
Glycoproteins/*genetics/metabolism
Hydrogen Peroxide/pharmacology
Male
Proto-Oncogene Proteins c-bcl-2/*genetics/metabolism
RNA, Messenger/genetics/metabolism
Real-Time Polymerase Chain Reaction/veterinary
Caspase 3
Glycoproteins
Proto-Oncogene Proteins c-bcl-2
RNA, Messenger
Hydrogen Peroxide
Figure
-
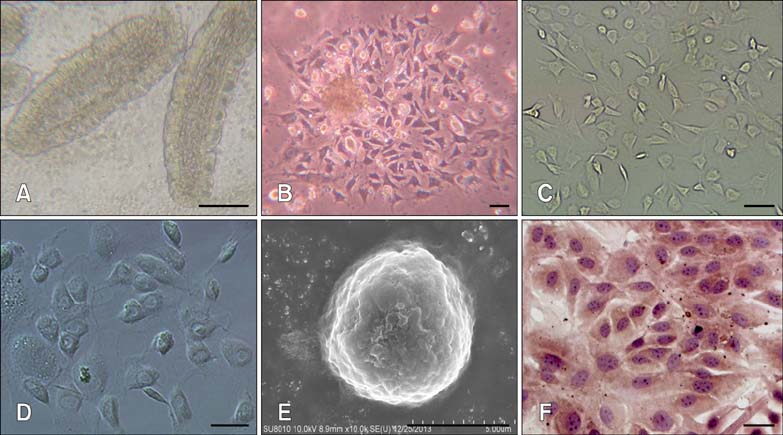
Fig. 1 Morphology of intestinal cells in culture. Phase contrast microscopy was performed to observed characteristics associated with the evolution of bovine IECs in primary cultures starting from adherent organoid structures (A) that gave rise to lager proliferating foci (B) which joined together to form a confluent monolayer. After purification, the subcultured epithelial cells grew dispersedly in the flask (C). The individual epithelial cell had a dome-like and rounded morphology when viewed with a differential interference contrast microscope (D) or scanning electron microscope (E), respectively. The nuclei and cytoplasm of the hematoxylin- and eosin-stained cells were blue and pink, respectively (F). ×40 (A), ×200 (B ~ D, and F), and ×15,000 (E). Scale bars = 200 µm (A) or 50 µm (B ~ F).
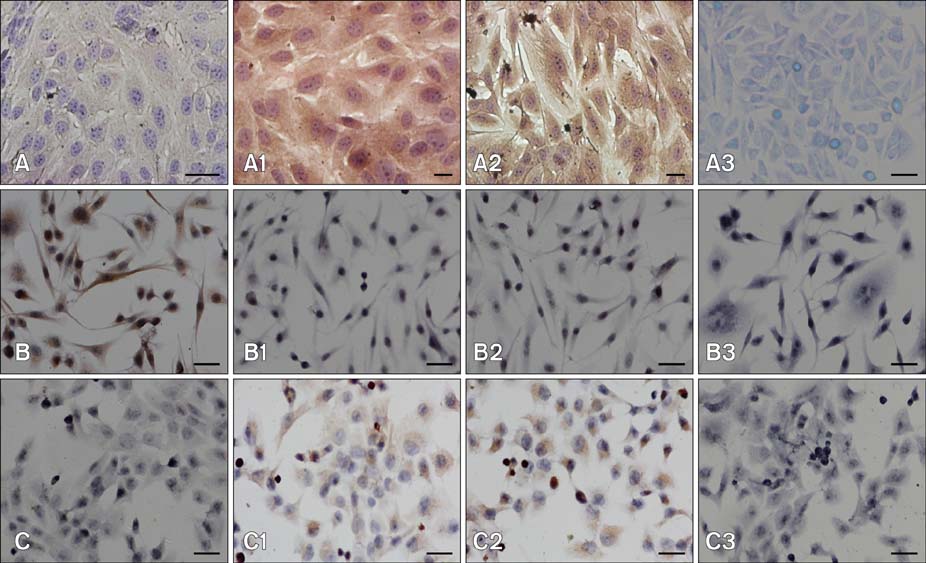
Fig. 2 Characterization of cells stained for immunocytochemistry. Panels show the primary cultures, V79, and IEC-6 cells, and indicate cells stained with antibodies against vimentin, PCK, CK18, or PBS (as the negative control), from left to right, respectively. Note that both the primary cultures (A1 and 2) and IEC-6 cells (C1 and 2) were stained yellow by the antibodies against PCK and CK18, but not by the anti-vimentin antibody (A and C). In contrast, the V79 cells displayed opposite patterns of immunoreactivity (B ~ B2). ×400 (A1 and 2), ×200 (A, A3, B ~ B3, and C ~ C3). Scale bars = 20 µm (A1 and 2) or 50 µm (A, A3, B ~ B3, and C ~ C3).
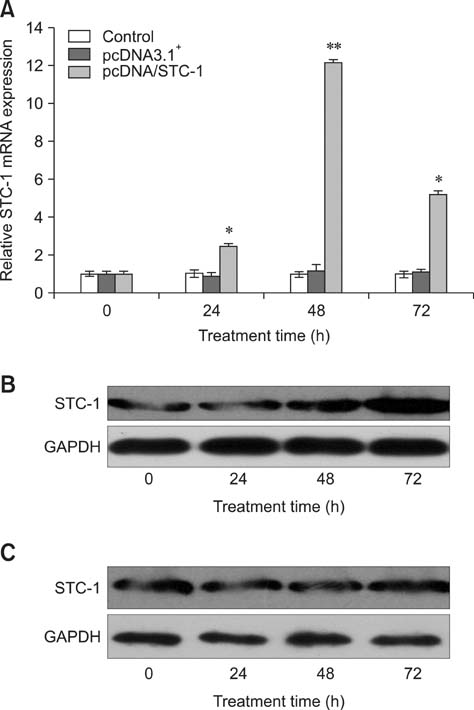
Fig. 3 Analysis of STC-1 mRNA and protein expression in the transfected IECs. The expression of STC-1 mNRA in cells transfected with the pcDNA/STC-1 construct was maximized approximately 48 h after transfection as measured by real-time PCR (gray column in A; *p < 0.01 or **p < 0.001 vs. the control cells). Protein expression was maximized at 72 h as measured by Western blotting (B). In contrast, no significant changes were observed in STC-1 mRNA or protein expression in cells transfected with the pcDNA3.1+ vector (black column in A and C).
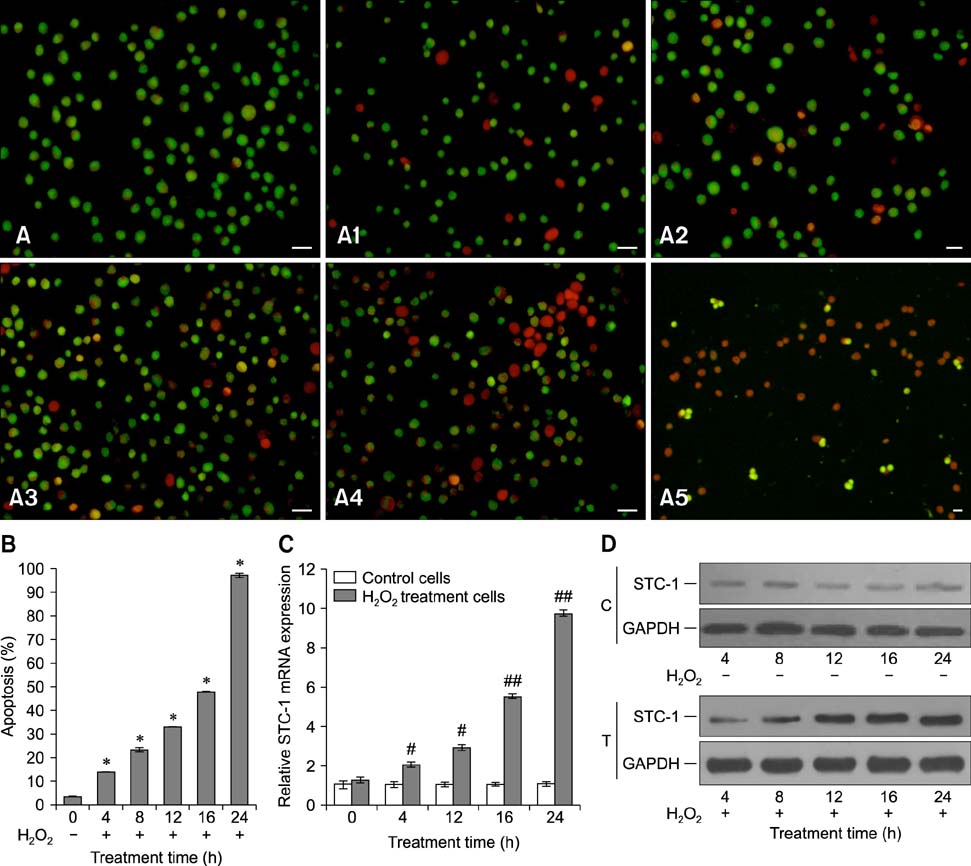
Fig. 4 Detection of viability, apoptosis, and STC-1 expression of transfected IECs. A time-dependent decrease in cell viability (A ~ A5, cells treated with H2O2 for 0, 4, 8, 12, 16, and 24 h) and survival rate (B, *p < 0.001 vs. 0 h) was observed for cells treated with H2O2 using AO/EB double staining and a trypan blue exclusion assay, respectively. Additionally, time-dependent increases in STC-1 mRNA (C, #p < 0.01 or ##p < 0.001 vs. the control cells) and protein expression (D, C: control cells, T: H2O2-treated cells) were found in the cells exposed to 200 µM H2O2 using real-time PCR and Western blotting, respectively. ×400 (A ~ A5). Scale bars = 20 µm (A ~ A5).
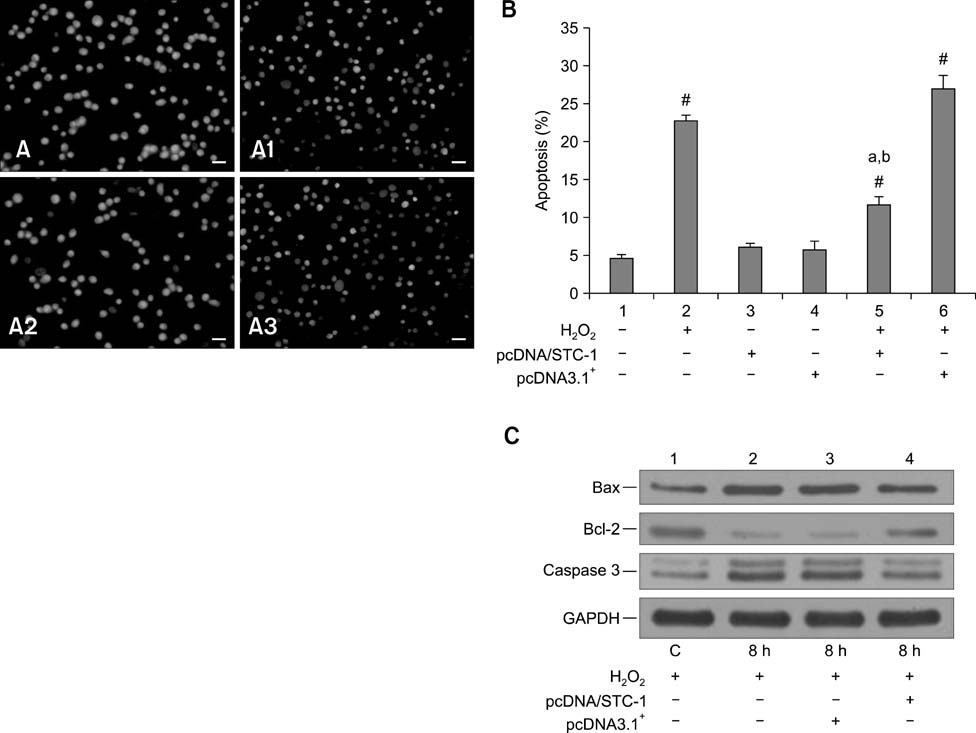
Fig. 5 The effects of STC-1 over-expression on the viability, apoptosis rate, and apoptosis-related protein expression in IECs. (A ~ A3) Viability of cells grown normally, treated with H2O2 for 8 h, or pre-transfected with pcDNA/STC-1 or pcDNA3.1+ for 48 h before H2O2 treatment, respectively. In cells that over-overexpressed STC-1, H2O2-induced cellular injury was significantly attenuated as measured by AO/EB double staining (A3). (B) A significant decrease of apoptosis induced by H2O2 was observed in cells transfected with pcDNA/STC-1 (fifth column) as measured by a trypan blue exclusion assay (#p < 0.01 vs. the control; ap < 0.01 vs. cells treated with H2O2 alone; bp < 0.01 vs. the pcDNA3.1+-transfected cells treated with H2O2). (C) Expression of apoptosis-related factors measured by Western blotting. Lane 1, Cells grown normally as the control; Lane 2, cells treated with H2O2 alone for 8 h; Lane 3, cells treated with H2O2 alone for 8 h after transfection with pcDNA3.1+ for 48 h; Lane 4, cells treated with H2O2 alone for 8 h after transfection with pcDNA/STC-1 for 48 h. Cells transfected with pcDNA/STC-1 showed a significant up-regulation of Bcl-2 expression and a slight down-regulation of caspase 3 expression. ×400 (A1~A4). Scale bars = 20 µm (A1 ~ A4).
Reference
-
1. Arigami T, Uenosono Y, Ishigami S, Hagihara T, Haraguchi N, Matsushita D, Yanagita S, Nakajo A, Okumura H, Hokita S, Natsugoe S. Expression of stanniocalcin 1 as a potential biomarker of gastric cancer. Oncology. 2012; 83:158–164.
Article2. Bae JY, Ahn SJ, Han W, Noh DY. Peroxiredoxin I and II inhibit H2O2-induced cell death in MCF-7 cell lines. J Cell Biochem. 2007; 101:1038–1045.
Article3. Baioni L, Basini G, Bussolati S, Grasselli F. Stanniocalcin 1 affects redox status of swine granulosa cells. Regul Pept. 2011; 168:45–49.
Article4. Bartsch H, Nair J. Chronic inflammation and oxidative stress in the genesis and perpetuation of cancer: role of lipid peroxidation, DNA damage, and repair. Langenbecks Arch Surg. 2006; 391:499–510.
Article5. Chang ACM, Janosi J, Hulsbeek M, de Jong D, Jeffrey KJ, Noble JR, Reddel RR. A novel human cDNA highly homologous to the fish hormone stanniocalcin. Mol Cell Endocrinol. 1995; 112:241–247.
Article6. De Niu P, Radman DP, Jaworski EM, Deol H, Gentz R, Su J, Olsen HS, Wagner GF. Development of a human stanniocalcin radioimmunoassay: serum and tissue hormone levels and pharmacokinetics in the rat. Mol Cell Endocrinol. 2000; 162:131–144.
Article7. Deol H, Stasko SE, De Niu P, James KA, Wagner GF. Post-natal ontogeny of stanniocalcin gene expression in rodent kidney and regulation by dietary calcium and phosphate. Kidney Int. 2001; 60:2142–2152.
Article8. Föllmann W, Weber S, Birkner S. Primary cell cultures of bovine colon epithelium: isolation and cell culture of colonocytes. Toxicol In Vitro. 2000; 14:435–445.
Article9. Feghali CA, Wright TM. Cytokines in acute and chronic inflammation. Front Biosci. 1997; 2:d12–d26.
Article10. Freshney RI, Freshney MG. Culture of Epithelial Cells. 2nd ed. New York: Wiley-Liss;2004. p. 303–336.11. Huang L, Belousova T, Chen M, DiMattia G, Liu D, Sheikh-Hamad D. Overexpression of stanniocalcin-1 inhibits reactive oxygen species and renal ischemia/reperfusion injury in mice. Kidney Int. 2012; 82:867–877.
Article12. Huang L, Garcia G, Lou Y, Zhou Q, Truong LD, DiMattia G, Lan XR, Lan HY, Wang Y, Sheikh-Hamad D. Anti-inflammatory and renal protective actions of stanniocalcin-1 in a model of anti-glomerular basement membrane glomerulonephritis. Am J Pathol. 2009; 174:1368–1378.
Article13. Kopa Z, Wenzel J, Papp GK, Haidl G. Role of granulocyte elastase and interleukin-6 in the diagnosis of male genital tract inflammation. Andrologia. 2005; 37:188–194.
Article14. La Ragione RM, Cooley WA, Woodward MJ. The role of fimbriae and flagella in the adherence of avian strains of Escherichia coli O78:K80 to tissue culture cells and tracheal and gut explants. J Med Microbiol. 2000; 49:327–338.
Article15. Law AYS, Ching LY, Lai KP, Wong CKC. Identification and characterization of the hypoxia-responsive element in human stanniocalcin-1 gene. Mol Cell Endocrinol. 2010; 314:118–127.
Article16. Lee H, Stabel J, Kehrli ME Jr. Cytokine gene expression in ileal tissues of cattle infected with Mycobacterium paratuberculosis. Vet Immunol Immunopathol. 2001; 82:73–85.
Article17. Liu G, Yang G, Chang B, Mercado-Uribe I, Huang M, Zheng J, Bast RC, Lin SH, Liu J. Stanniocalcin 1 and ovarian tumorigenesis. J Natl Cancer Inst. 2010; 102:812–827.
Article18. Lykkesfeldt J, Svendsen O. Oxidants and antioxidants in disease: oxidative stress in farm animals. Vet J. 2007; 173:502–511.
Article19. McGill G, Shimamura A, Bates RC, Savage RE, Fisher DE. Loss of matrix adhesion triggers rapid transformationselective apoptosis in fibroblasts. J Cell Biol. 1997; 138:901–911.
Article20. Moyer MP, Aust JB. Human colon cells: culture and in vitro transformation. Science. 1984; 224:1445–1447.
Article21. Serlachius M, Andersson LC. Upregulated expression of stanniocalcin-1 during adipogenesis. Exp Cell Res. 2004; 296:256–264.
Article22. Sheikh-Hamad D. Mammalian stanniocalcin-1 activates mitochondrial antioxidant pathways: new paradigms for regulation of macrophages and endothelium. Am J Physiol Renal Physiol. 2010; 298:F248–F254.
Article23. Tremblay G, Delbecchi L, Loiselle MC, Ster C, Wagner GF, Talbot BG, Lacasse P. Serum levels of stanniocalcin-1 in Holstein heifers and cows. Domest Anim Endocrinol. 2009; 36:105–109.
Article24. Wagner GF, Jaworski E. Calcium regulates stanniocalcin mRNA levels in primary cultured rainbow trout corpuscles of stannius. Mol Cell Endocrinol. 1994; 99:315–322.
Article25. Wagner GF, Hampong M, Park CM, Copp DH. Purification, characterization, and bioassay of teleocalcin, a glycoprotein from salmon corpuscles of Stannius. Gen Comp Endocrinol. 1986; 63:481–491.
Article26. Watson PR, Galyov EE, Paulin SM, Jones PW, Wallis TS. Mutation of invH, but Not stn, reduces Salmonella-induced enteritis in cattle. Infect immun. 1998; 66:1432–1438.
Article27. Wiseman H, Halliwell B. Damage to DNA by reactive oxygen and nitrogen species: role in inflammatory disease and progression to cancer. Biochem J. 1996; 313:17–29.
Article28. Wu L, Xi Z, Guo R, Liu S, Yang S, Liu D, Dong S, Guo D. Exogenous ARC down-regulates caspase-3 expression and inhibits apoptosis of broiler chicken cardiomyocytes exposed to hydrogen peroxide. Avian Pathol. 2013; 42:32–37.
Article29. Yeung HY, Lai KP, Chan HY, Mak NK, Wagner GF, Wong CKC. Hypoxia-inducible factor-1-mediated activation of stanniocalcin-1 in human cancer cells. Endocrinology. 2005; 146:4951–4960.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Paeoniflorin Protects Retinal Pigment Epithelial Cells from High Glucose-Induced Oxidative Damage by Activating Nrf2-Mediated HO-1 Signaling
- Erratum to "Morroniside Protects C2C12 Myoblasts from Oxidative Damage Caused by ROS-mediated Mitochondrial Damage and Induction of Endoplasmic Reticulum Stress" [Biomol Ther 32(3), 349-360 (2024)]
- Erratum to "Morroniside Protects C2C12 Myoblasts from Oxidative Damage Caused by ROS-mediated Mitochondrial Damage and Induction of Endoplasmic Reticulum Stress" [Biomol Ther 32(3), 349-360 (2024)]
- Carbon monoxide releasing molecule-2 protects mice against acute kidney injury through inhibition of ER stress
- Epilepsy and Oxidative Stress
