Vitamin D maintains E-cadherin intercellular junctions by downregulating MMP-9 production in human gingival keratinocytes treated by TNF-α
- Affiliations
-
- 1Laboratory for the Study of Molecular Biointerfaces, Department of Oral Histology and Developmental Biology, Program of Cell and Developmental Biology, School of Dentistry and Dental Research Institute, Seoul National University, Seoul, Korea. hyunmkim@snu.ac.kr
- KMID: 2461018
- DOI: http://doi.org/10.5051/jpis.2019.49.5.270
Abstract
- PURPOSE
Despite the well-known anti-inflammatory effects of vitamin D in periodontal health, its mechanism has not been fully elucidated. In the present study, the effect of vitamin D on strengthening E-cadherin junctions (ECJs) was explored in human gingival keratinocytes (HGKs). ECJs are the major type of intercellular junction within the junctional epithelium, where loose intercellular junctions develop and microbial invasion primarily occurs.
METHODS
HOK-16B cells, an immortalized normal human gingival cell line, were used for the study. To mimic the inflammatory environment, cells were treated with tumor necrosis factor-alpha (TNF-α). Matrix metalloproteinases (MMPs) in the culture medium were assessed by an MMP antibody microarray and gelatin zymography. The expression of various molecules was investigated using western blotting. The extent of ECJ development was evaluated by comparing the average relative extent of the ECJs around the periphery of each cell after immunocytochemical E-cadherin staining. Vitamin D receptor (VDR) expression was examined via immunohistochemical analysis.
RESULTS
TNF-α downregulated the development of the ECJs of the HGKs. Dissociation of the ECJs by TNF-α was accompanied by the upregulation of MMP-9 production and suppressed by a specific MMP-9 inhibitor, Bay 11-7082. Exogenous MMP-9 decreased the development of ECJs. Vitamin D reduced the production of MMP-9 and attenuated the breakdown of ECJs in the HGKs treated with TNF-α. In addition, vitamin D downregulated TNF-α-induced nuclear factor kappa B (NF-κB) signaling in the HGKs. VDR was expressed in the gingival epithelium, including the junctional epithelium.
CONCLUSIONS
These results suggest that vitamin D may avert TNF-α-induced downregulation of the development of ECJs in HGKs by decreasing the production of MMP-9, which was upregulated by TNF-α. Vitamin D may reinforce ECJs by downregulating NF-κB signaling, which is upregulated by TNF-α. Strengthening the epithelial barrier may be a way for vitamin D to protect the periodontium from bacterial invasion.
Keyword
MeSH Terms
-
Bays
Blotting, Western
Cadherins*
Cell Line
Down-Regulation
Epithelial Attachment
Epithelium
Gelatin
Humans*
Intercellular Junctions*
Keratinocytes*
Matrix Metalloproteinase 9
Matrix Metalloproteinases
NF-kappa B
Periodontium
Receptors, Calcitriol
Tumor Necrosis Factor-alpha
Up-Regulation
Vitamin D*
Vitamins*
Cadherins
Gelatin
Matrix Metalloproteinase 9
Matrix Metalloproteinases
NF-kappa B
Receptors, Calcitriol
Tumor Necrosis Factor-alpha
Vitamin D
Vitamins
Figure
-
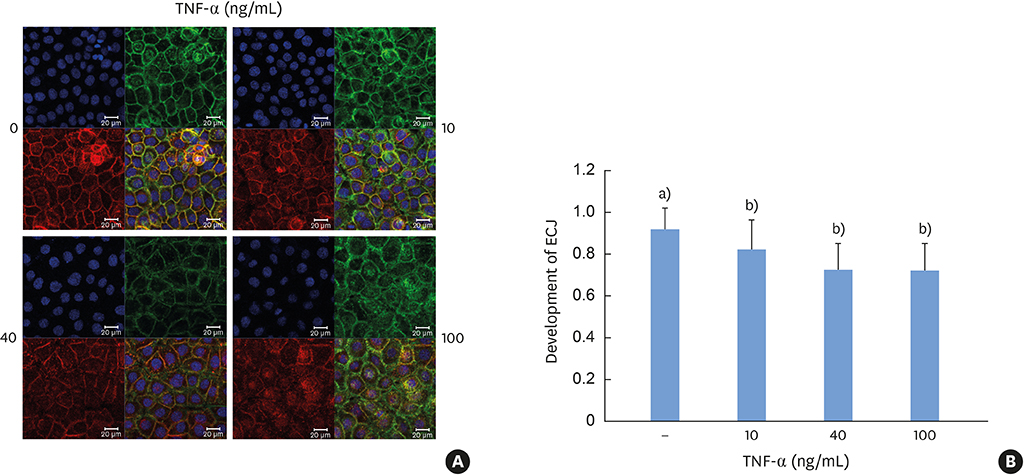
Figure 1 TNF-α suppresses the development of ECJs in HGKs in vitro. Cells were cultured for 24 hours for the establishment of the ECJs and then treated with various amounts of TNF-α (10, 40, 100 ng/mL). (A) ECJ development was detected by immunocytochemical staining of E-cadherin expression (red). In addition, F-actin (green) was stained with FITC-phalloidin and nuclei were stained with DAPI (blue). The lower right pictures in each set of pictures are the merged images of the E-cadherin, F-actin, and nuclei images.(B) The average ratio of the development of ECJs per cell was calculated. TNF-α: tumor necrosis factor-alpha, ECJ: E-cadherin junction, HGK: human gingival keratinocyte, DAPI: 4′,6-diamidino-2-phenylindole dihydrochloride. a)P<0.05 versus b)Nonparametric Kruskal-Wallis test and post hoc multiple comparison with the Dunn test.

Figure 2 Vitamin D suppresses the dissociation of the ECJs induced by TNF-α in HGKs in vitro. Cells were cultured for 24 hours for the establishment of ECJs and pretreated with vitamin D (10, 100 nM) or without it for an additional 24 hours. Then, cells were cultured with or without TNF-α for 24 hours along with vitamin D. (A) ECJ development was detected by immunocytochemical staining of E-cadherin expression (red). In addition, F-actin (green) was stained with FITC-phalloidin and nuclei were stained with DAPI (blue). The lower right pictures in each set of pictures are the merged images of the E-cadherin, F-actin, and nuclei images. (B) The average ratio of the development of ECJs per cell was calculated. ECJ: E-cadherin junction, TNF-α: tumor necrosis factor-alpha, HGK: human gingival keratinocyte, DAPI: 4′,6-diamidino-2-phenylindole dihydrochloride. a)P<0.05 versus b)Nonparametric Kruskal-Wallis test and post hoc multiple comparison with the Dunn test.

Figure 3 TIMP-1 suppresses the dissociation of ECJs induced by TNF-α in HGKs in vitro. Cells were cultured for 24 hours for the establishment of the ECJs and then treated with TNF-α (40 ng/mL), with or without TIMP-1 (20 ng/mL) for an additional 24 hours. (A) ECJ development was followed by immunocytochemical staining of E-cadherin expression (red). In addition, F-actin (green) was stained with FITC-phalloidin and nuclei were stained with DAPI (blue). The lower right pictures in each set of pictures are the merged images of the E-cadherin, F-actin, and nuclei images. (B) The average ratio of the development of ECJs per cell was calculated by taking the mean of the values that were obtained by dividing the length of the E-cadherin expressed at cell junctions by the total perimeter of each cell. TIMP-1: tissue inhibitor of metalloproteinase-1, ECJ: E-cadherin junction, TNF-α: tumor necrosis factor-alpha, HGK: human gingival keratinocyte, DAPI: 4′,6-diamidino-2-phenylindole dihydrochloride. a)P<0.05 versus b)Nonparametric Kruskal-Wallis test and post hoc multiple comparison with the Dunn test.

Figure 4 TNF-α upregulates the secretion of MMP-9, as shown by antibody microarray analysis, and gelatinolytic digestion by the secreted MMP-9, as demonstrated by gelatin zymography in HGKs in vitro, respectively. (A) Cells were cultured for 24 hours for the establishment of the ECJs and then treated with TNF-α (40 ng/mL). Antibody microarray analysis of MMPs was performed by applying conditioned culture medium to the antibody microarray chip for MMPs. The table contains a list of the MMPs or TIMPs in the array. The numbers under the MMP-9 blots indicate the intensity of MMP-9 standardized using the positive controls as references (boxes). (B) Cells were cultured for 24 hours for the establishment of the ECJs and then treated with various amounts of TNF-α (10, 40, 100 ng/mL). Secretion of MMP-9 into the culture medium was assessed by gelatin zymogram analysis. The numbers under the MMP-9 bands indicate the activity of MMP-9. TNF-α: tumor necrosis factor-alpha, MMP-9: matrix metalloproteinase-9, HGK: human gingival keratinocyte, ECJ: E-cadherin junction, Pos: positive control for the chip reactions, Neg: negative control for the chip reactions.

Figure 5 A specific inhibitor of MMP-9 reverses the effect of TNF-α on the development of ECJs in HGKs in vitro. Cells were cultured for 24 hours for the establishment of the ECJs and then treated with TNF-α (40 ng/mL) along with various amounts of an MMP-9 inhibitor (5, 20, or 50 µM) or without it for an additional 24 hours. (A) ECJ development was followed by immunocytochemical staining of E-cadherin expression (red). In addition, F-actin (green) was stained with FITC-phalloidin and nuclei were stained with DAPI (blue). The lower right pictures in each set of pictures are the merged images of the E-cadherin, F-actin, and nuclei images. (B) The average extent of the development of ECJs per cell was calculated by taking the mean of the values that were obtained by dividing the length of the E-cadherin expressed at cell junctions by the total perimeter of each cell. MMP-9: matrix metalloproteinase-9, TNF-α: tumor necrosis factor-alpha, ECJ: E-cadherin junction, HGK: human gingival keratinocyte, DAPI: 4′,6-diamidino-2-phenylindole dihydrochloride. a)P<0.05 versus b)Nonparametric Kruskal-Wallis test and post hoc multiple comparison with the Dunn test.
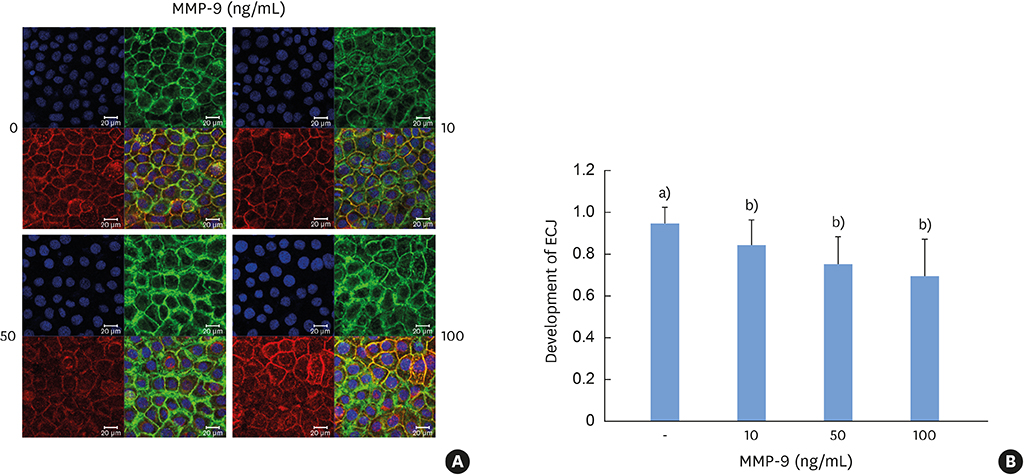
Figure 6 MMP-9 induces the dissociation of ECJs in HGKs in vitro. Cells were cultured for 24 hours for the establishment of the ECJs and then treated with various amounts of MMP-9 (10, 50, 100 ng/mL). (A) ECJ development was followed by immunocytochemical staining of E-cadherin expression (red). In addition, F-actin (green) was stained with FITC-phalloidin and nuclei were stained with DAPI (blue). The lower right pictures in each set of pictures are the merged images of the E-cadherin, F-actin, and nuclei images. (B) The average extent of the development of ECJs per cell was calculated by taking the mean of the values that were obtained by dividing the length of the E-cadherin expressed at cell junctions by the total perimeter of each cell. MMP-9: matrix metalloproteinase-9, ECJ: E-cadherin junction, HGK: human gingival keratinocyte, DAPI: 4′,6-diamidino-2-phenylindole dihydrochloride. a)P<0.05 versus b)Nonparametric Kruskal-Wallis test and post hoc multiple comparison with the Dunn test.

Figure 7 Vitamin D downregulates the TNF-α-induced secretion of MMP-9 and the gelatinolytic digestion by the MMP-9 upregulated by TNF-α in HGKs in vitro. Cells were cultured for 24 hours for the establishment of ECJs and pretreated with or without vitamin D (10 or 100 nM) for an additional 24 hours. Then, cells were cultured with or without TNF-α (40 ng/mL) along with vitamin D for 24 and 48 hours (A) or 48 hours (B). (A) Antibody microarray analysis of MMPs was performed by applying conditioned culture medium to the antibody microarray of MMPs. Details on the arrays are shown in the legend of Figure 4A. The numbers under the MMP-9 blots indicate the intensity of MMP-9 standardized using the positive controls as references (boxes). (B) Secretion of MMP-9 into the culture medium was assessed by gelatin zymogram analysis. The numbers under the MMP-9 bands indicate the activity of MMP-9. TNF-α: tumor necrosis factor-alpha, MMP-9: matrix metalloproteinase-9, HGK: human gingival keratinocyte, ECJ: E-cadherin junction.

Figure 8 NF-κB inhibition by Bay 11-7082 reverses the TNF-α-induced effects on HGKs in vitro. Cells were cultured for 24 hours for the establishment of ECJs and then treated with TNF-α (40 ng/mL) along with Bay 11-7082 (Bay11; 10 nM) or without it for an additional 24 hours. (A) ECJ development was followed by immunocytochemical staining of E-cadherin expression (red). In addition, F-actin (green) was stained with FITC-phalloidin and nuclei were stained with DAPI (blue). The lower right pictures in each set of pictures are the merged images of the E-cadherin, F-actin, and nuclei images. (B) The average ratio of the development of ECJs per cell was calculated by taking the mean of the values which were obtained by dividing the length of the E-cadherin expressed at cell junctions by the total perimeter of each cell. NF-κB: nuclear factor kappa B, TNF-α: tumor necrosis factor-alpha, HGK: human gingival keratinocyte, ECJ: E-cadherin junction, DAPI: 4′,6-diamidino-2-phenylindole dihydrochloride. a)P<0.05 versus b)Nonparametric Kruskal-Wallis test and post hoc multiple comparison with the Dunn test.
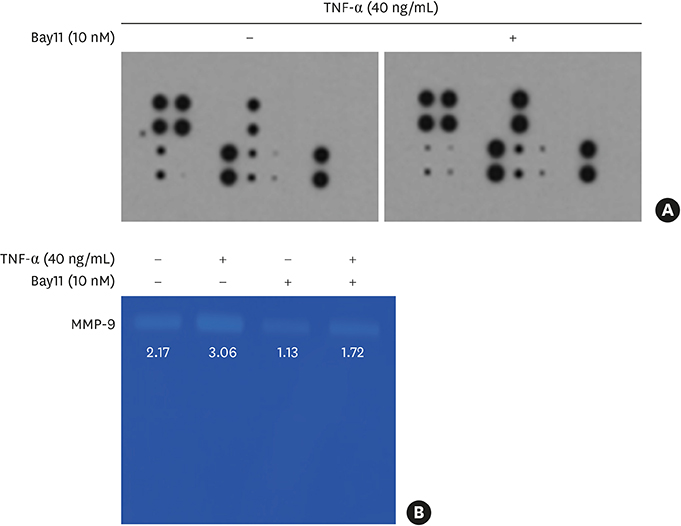
Figure 9 The inhibition of NF-κB signaling by Bay 11-7082 lowers the secretion of MMP-9 into the culture medium and the gelatinolytic digestion by the secreted MMP-9, upregulated by TNF-α. Cells were cultured for 24 hours for the establishment of ECJs and then treated with TNF-α (40 ng/mL) along with Bay 11-7082 (Bay11; 10 nM) or without it for an additional 24 hours. (A) Antibody microarray analyses of MMPs were performed by applying culture medium to the MMPs antibody microarray chip. Representative data from the triplicate measurements are shown. Details on the arrays are shown in the legend of Figure 4A. The numbers under the MMP-9 blots indicate the intensity of MMP-9 standardized using the positive controls as references (boxes). (B) Secretion of MMP-9 into the culture medium was assessed by gelatin zymogram analysis. The numbers under the MMP-9 bands indicate the activity of MMP-9. NF-κB: nuclear factor kappa B, MMP-9: matrix metalloproteinase-9, TNF-α: tumor necrosis factor-alpha, ECJ: E-cadherin junction.

Figure 10 Vitamin D inhibits TNF-α-induced NF-κB signaling, as shown by western blotting. Cells were cultured for 48 hours and then pretreated with vitamin D (10, 100, or 1,000 nM) or without it for 24 hours. Before cell lysis for western blotting, cells were treated with TNF-α (100 ng/mL) for 10 minutes. Bay 11-7082 (Bay 11; 1 nM), a pharmacological inactivator of NF-κB signaling, was used as an internal control of the immunoblotting analyses to show that TNF-α-induced NF-κB signaling was reduced when NF-κB signaling was pharmacologically inactivated in HGKs. TNF-α: tumor necrosis factor-alpha, NF-κB: nuclear factor kappa B, HGK: human gingival keratinocyte, GAPDH: glyceraldehyde 3-phosphate dehydrogenase.
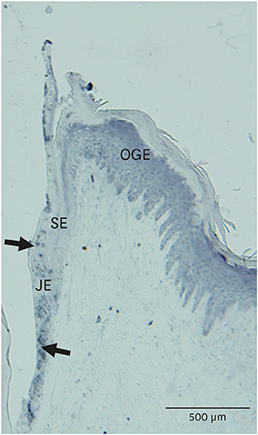
Figure 11 VDR is expressed in the gingival epithelium. VDR expression in gingival epithelium surrounding rat mandibular molars was examined using immunohistochemical methods. Expression of VDR was apparent in the JE (arrows). Expression of VDR was also localized in the OGE and SE. VDR: vitamin D receptor, JE: junctional epithelium, OGE: oral gingival epithelium, SE: sulcular epithelium.
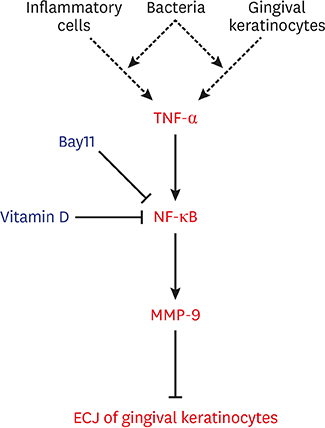
Figure 12 Diagram of a hypothetical molecular mechanism by which vitamin D suppresses the dissociation of ECJs induced by TNF-α. Vitamin D suppresses the NF-κB activation induced by TNF-α, which abolishes the detrimental action of TNF-α through downregulating the production of MMP-9, which breaks the integrity of ECJs. TNF-α may be produced by gingival keratinocytes infected with bacteria in an autocrine fashion at the early epithelial stage of infection [7] or could be externally provided from inflammatory cells in connective tissue at the late stage of inflammation. In summary, the present study suggests that vitamin D may be protective for periodontal health by strengthening the epithelial barrier. ECJ: E-cadherin junction, TNF-α: tumor necrosis factor-alpha, NF-κB: nuclear factor kappa B, MMP-9: matrix metalloproteinase-9.
Reference
-
1. Alshouibi EN, Kaye EK, Cabral HJ, Leone CW, Garcia RI. Vitamin D and periodontal health in older men. J Dent Res. 2013; 92:689–693.
Article2. Lin Z, Li W. The roles of vitamin D and its analogs in inflammatory diseases. Curr Top Med Chem. 2016; 16:1242–1261.
Article3. Garcia MN, Hildebolt CF, Miley DD, Dixon DA, Couture RA, Spearie CL, et al. One-year effects of vitamin D and calcium supplementation on chronic periodontitis. J Periodontol. 2011; 82:25–32.
Article4. Dietrich T, Nunn M, Dawson-Hughes B, Bischoff-Ferrari HA. Association between serum concentrations of 25-hydroxyvitamin D and gingival inflammation. Am J Clin Nutr. 2005; 82:575–580.
Article5. Dietrich T, Joshipura KJ, Dawson-Hughes B, Bischoff-Ferrari HA. Association between serum concentrations of 25-hydroxyvitamin D3 and periodontal disease in the US population. Am J Clin Nutr. 2004; 80:108–113.6. Hiremath VP, Rao CB, Naik V, Prasad KV. Anti-inflammatory effect of vitamin D on gingivitis: a dose-response randomised control trial. Oral Health Prev Dent. 2013; 11:61–69.7. De Filippis A, Fiorentino M, Guida L, Annunziata M, Nastri L, Rizzo A. Vitamin D reduces the inflammatory response by Porphyromonas gingivalis infection by modulating human β-defensin-3 in human gingival epithelium and periodontal ligament cells. Int Immunopharmacol. 2017; 47:106–117.
Article8. McMahon L, Schwartz K, Yilmaz O, Brown E, Ryan LK, Diamond G. Vitamin D-mediated induction of innate immunity in gingival epithelial cells. Infect Immun. 2011; 79:2250–2256.
Article9. Bosshardt DD, Lang NP. The junctional epithelium: from health to disease. J Dent Res. 2005; 84:9–20.
Article10. Damek-Poprawa M, Korostoff J, Gill R, DiRienzo JM. Cell junction remodeling in gingival tissue exposed to a microbial toxin. J Dent Res. 2013; 92:518–523.
Article11. Hatakeyama S, Yaegashi T, Oikawa Y, Fujiwara H, Mikami T, Takeda Y, et al. Expression pattern of adhesion molecules in junctional epithelium differs from that in other gingival epithelia. J Periodontal Res. 2006; 41:322–328.
Article12. Bosshardt DD. The periodontal pocket: pathogenesis, histopathology and consequences. Periodontol 2000. 2018; 76:43–50.
Article13. Lee G, Kim HJ, Kim HM. RhoA-JNK regulates the E-cadherin junctions of human gingival epithelial cells. J Dent Res. 2016; 95:284–291.
Article14. Lewis JE, Wahl JK 3rd, Sass KM, Jensen PJ, Johnson KR, Wheelock MJ. Cross-talk between adherens junctions and desmosomes depends on plakoglobin. J Cell Biol. 1997; 136:919–934.
Article15. Ye P, Chapple CC, Kumar RK, Hunter N. Expression patterns of E-cadherin, involucrin, and connexin gap junction proteins in the lining epithelia of inflamed gingiva. J Pathol. 2000; 192:58–66.
Article16. Miyagawa T, Fujita T, Yumoto H, Yoshimoto T, Kajiya M, Ouhara K, et al. Azithromycin recovers reductions in barrier function in human gingival epithelial cells stimulated with tumor necrosis factor-α. Arch Oral Biol. 2016; 62:64–69.
Article17. Choi YS, Baek K, Choi Y. Estrogen reinforces barrier formation and protects against tumor necrosis factor alpha-induced barrier dysfunction in oral epithelial cells. J Periodontal Implant Sci. 2018; 48:284–294.
Article18. Park NH, Min BM, Li SL, Huang MZ, Cherick HM, Doniger J. Immortalization of normal human oral keratinocytes with type 16 human papillomavirus. Carcinogenesis. 1991; 12:1627–1631.
Article19. Jin C, Lee G, Oh C, Kim HJ, Kim HM. Substrate roughness induces the development of defective E-cadherin junctions in human gingival keratinocytes. J Periodontal Implant Sci. 2017; 47:116–131.
Article20. Symowicz J, Adley BP, Gleason KJ, Johnson JJ, Ghosh S, Fishman DA, et al. Engagement of collagen-binding integrins promotes matrix metalloproteinase-9-dependent E-cadherin ectodomain shedding in ovarian carcinoma cells. Cancer Res. 2007; 67:2030–2039.
Article21. Zheng G, Lyons JG, Tan TK, Wang Y, Hsu TT, Min D, et al. Disruption of E-cadherin by matrix metalloproteinase directly mediates epithelial-mesenchymal transition downstream of transforming growth factor-beta1 in renal tubular epithelial cells. Am J Pathol. 2009; 175:580–591.
Article22. Koli K, Keski-Oja J. 1α,25-dihydroxyvitamin D3 and its analogues down-regulate cell invasion-associated proteases in cultured malignant cells. Cell Growth Differ. 2000; 11:221–229.23. Bao BY, Yeh SD, Lee YF. 1α,25-dihydroxyvitamin D3 inhibits prostate cancer cell invasion via modulation of selective proteases. Carcinogenesis. 2006; 27:32–42.
Article24. Chiang KC, Yeh CN, Hsu JT, Jan YY, Chen LW, Kuo SF, et al. The vitamin D analog, MART-10, represses metastasis potential via downregulation of epithelial-mesenchymal transition in pancreatic cancer cells. Cancer Lett. 2014; 354:235–244.
Article25. Liu T, Zhang L, Joo D, Sun SC. NF-κB signaling in inflammation. Signal Transduct Target Ther. 2017; 2:17023.
Article26. Yuan FN, Valiyaparambil J, Woods MC, Tran H, Pant R, Adams JS, et al. Vitamin D signaling regulates oral keratinocyte proliferation in vitro and in vivo. Int J Oncol. 2014; 44:1625–1633.
Article27. Fischer KD, Agrawal DK. Vitamin D regulating TGF-β induced epithelial-mesenchymal transition. Respir Res. 2014; 15:146.
Article28. Lopes N, Carvalho J, Durães C, Sousa B, Gomes M, Costa JL, et al. 1α,25-dihydroxyvitamin D3 induces de novo E-cadherin expression in triple-negative breast cancer cells by CDH1-promoter demethylation. Anticancer Res. 2012; 32:249–257.29. Larriba MJ, García de Herreros A, Muñoz A. Vitamin D and the epithelial to mesenchymal transition. Stem Cells Int. 2016; 2016:6213872.
Article30. Fujita T, Yumoto H, Shiba H, Ouhara K, Miyagawa T, Nagahara T, et al. Irsogladine maleate regulates epithelial barrier function in tumor necrosis factor-α-stimulated human gingival epithelial cells. J Periodontal Res. 2012; 47:55–61.
Article31. Halldorsson S, Gudjonsson T, Gottfredsson M, Singh PK, Gudmundsson GH, Baldursson O. Azithromycin maintains airway epithelial integrity during Pseudomonas aeruginosa infection. Am J Respir Cell Mol Biol. 2010; 42:62–68.
Article32. Carayol N, Campbell A, Vachier I, Mainprice B, Bousquet J, Godard P, et al. Modulation of cadherin and catenins expression by tumor necrosis factor-α and dexamethasone in human bronchial epithelial cells. Am J Respir Cell Mol Biol. 2002; 26:341–347.
Article33. Yi JY, Jung YJ, Choi SS, Chung E. TNF-alpha downregulates E-cadherin and sensitizes response to γ-irradiation in Caco-2 cells. Cancer Res Treat. 2009; 41:164–170.
Article34. Saito T, Yoshida K, Matsumoto K, Saeki K, Tanaka Y, Ong SM, et al. Inflammatory cytokines induce a reduction in E-cadherin expression and morphological changes in MDCK cells. Res Vet Sci. 2014; 96:288–291.
Article35. Vandooren J, Van den Steen PE, Opdenakker G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): the next decade. Crit Rev Biochem Mol Biol. 2013; 48:222–272.
Article36. Ji S, Choi Y. Point-of-care diagnosis of periodontitis using saliva: technically feasible but still a challenge. Front Cell Infect Microbiol. 2015; 5:65.
Article37. Kim HD, Shin MS, Kim HT, Kim MS, Ahn YB. Incipient periodontitis and salivary molecules among Korean adults: association and screening ability. J Clin Periodontol. 2016; 43:1032–1040.
Article38. Hayden MS, Ghosh S. Regulation of NF-κB by TNF family cytokines. Semin Immunol. 2014; 26:253–266.
Article39. Zhao B, Li R, Yang F, Yu F, Xu N, Zhang F, et al. LPS-induced vitamin D receptor decrease in oral keratinocytes is associated with oral lichen planus. Sci Rep. 2018; 8:763.
Article40. Janjetovic Z, Zmijewski MA, Tuckey RC, DeLeon DA, Nguyen MN, Pfeffer LM, et al. 20-Hydroxycholecalciferol, product of vitamin D3 hydroxylation by P450scc, decreases NF-κB activity by increasing IκBα levels in human keratinocytes. PLoS One. 2009; 4:e5988.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Substrate roughness induces the development of defective E-cadherin junctions in human gingival keratinocytes
- Effects of Interleukin 4 on the Production of Interleukin 6 in Human Keratinocytes
- The mRNA expression of MMP-1, TIMP-1, TGF-beta1 in gingival keratocytes from gingival hyperplasia induced by cyclosporine A
- The Expression of MT1-MMP and E-cadherin mRNA in Invasive Cervical Cancer
- The Effects of Calcium and Retinoic Acid on Epidermal Desmosomes
