Isomangiferin, a Novel Potent Vascular Endothelial Growth Factor Receptor 2 Kinase Inhibitor, Suppresses Breast Cancer Growth, Metastasis and Angiogenesis
- Affiliations
-
- 1Research Center of Basic Medical Sciences, School of Basic Medical Sciences, Hubei University of Science and Technology, Xianning, China. humeichun.530@163.com
- 2Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai, China.
- 3Cancer Center, Sanford Burnham Prebys Medical Discovery Institute, La Jolla, USA.
- 4Department of Optoelectronic Engineering, School of Electrical and Information Engineering, Hubei University of Science and Technology, Xianning, China.
- KMID: 2441863
- DOI: http://doi.org/10.4048/jbc.2018.21.1.11
Abstract
- PURPOSE
Vascular endothelial growth factor (VEGF) signal transduction mainly depends on its binding to VEGF receptor 2 (VEGFR-2). VEGF downstream signaling proteins mediate several of its effects in cancer progression, including those on tumor growth, metastasis, and blood vessel formation. The activation of VEGFR-2 signaling is a hallmark of and is considered a therapeutic target for breast cancer. Here, we report a study of the regulation of the VEGFR-2 signaling pathway by a small molecule, isomangiferin.
METHODS
A human breast cancer xenograft mouse model was used to investigate the efficacy of isomangiferin in vivo. The inhibitory effect of isomangiferin on breast cancer cells and the underlying mechanism were examined in vitro.
RESULTS
Isomangiferin suppressed tumor growth in xenografts. In vitro, isomangiferin treatment inhibited cancer cell proliferation, migration, invasion, and adhesion. The effect of isomangiferin on breast cancer growth was well coordinated with its suppression of angiogenesis. A rat aortic ring assay revealed that isomangiferin significantly inhibited blood vessel formation during VEGF-induced microvessel sprouting. Furthermore, isomangiferin treatment inhibited VEGF-induced proliferation of human umbilical vein endothelial cells and the formation of capillary-like structures. Mechanistically, isomangiferin induced caspase-dependent apoptosis of breast cancer cells. Furthermore, VEGF-induced activation of the VEGFR-2 kinase pathway was down-regulated by isomangiferin.
CONCLUSION
Our findings demonstrate that isomangiferin exerts anti-breast cancer effects via the functional inhibition of VEGFR-2. Pharmaceutically targeting VEGFR-2 by isomangiferin could be an effective therapeutic strategy for breast cancer.
Keyword
MeSH Terms
-
Angiogenesis Inhibitors
Animals
Apoptosis
Blood Vessels
Breast Neoplasms
Cell Proliferation
Heterografts
Human Umbilical Vein Endothelial Cells
Humans
In Vitro Techniques
Mice
Microvessels
Neoplasm Metastasis
Phosphotransferases
Rats
Receptors, Vascular Endothelial Growth Factor*
Signal Transduction
Vascular Endothelial Growth Factor A*
Vascular Endothelial Growth Factor Receptor-2*
Angiogenesis Inhibitors
Phosphotransferases
Receptors, Vascular Endothelial Growth Factor
Vascular Endothelial Growth Factor A
Vascular Endothelial Growth Factor Receptor-2
Figure
-
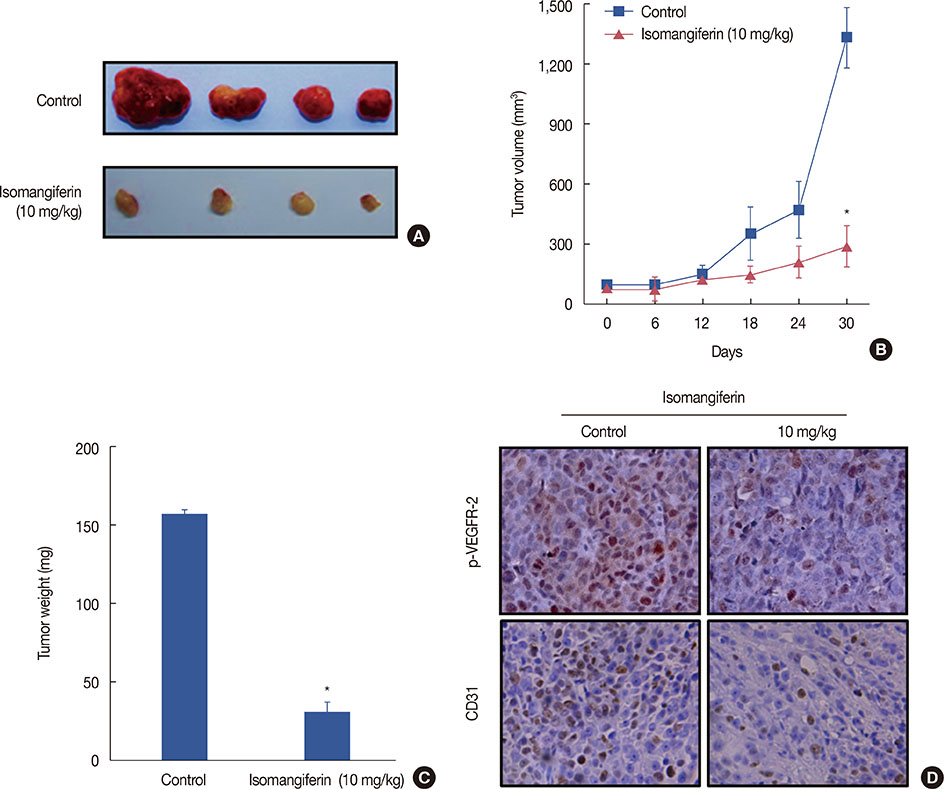
Figure 1 Isomangiferin inhibits breast tumor growth and vascular endothelial growth factor receptor 2 (VEGFR-2) signaling pathway in vivo. Human breast cancer cells MDA-MB-231 were injected subcutanously into 5-week-old BALB/cA nude mice (2×106 per mouse). When subcutanous tumors grew to about 100 mm3, the mice were intraperitoneally treated with or without isomangiferin (10 mg/kg/day). (A) Photos for isomangiferin treated or non-treated tumors. (B) Isomangiferin supressed MDA-MB-231 xenografts growth. Tumor volume was recorded every 6 days and the tumor growth curve was drafted through Graphpad Prism 5 software package. Values are shown as mean±standard error of the mean (SEM) of three independent experiments. (C) Isomangiferin inhibited breast tumor growth as measured by tumor weight. Values are shown as mean±SEM of three independent experiments. (D) Immunohistochemical staining revealed that isomangiferin inhibited VEGFR-2 signaling pathway by blotting phosphorylated VEGFR-2 (p-VEGFR-2) and reducing CD31 expression. Tumor sections from isomangiferin-treated and isomangiferin-untreated groups were stained using p-VEGFR-2 and CD31 antibodies, and the number of positive cells was counted (IHC stain for p-VEGFR-2, CD31, ×400). *p<0.01 vs. control.

Figure 2 Isomangiferin inhibits breast cancer cell proliferation, migration and invasion. (A) Isomangiferin inhibited different types of breast cancer cells proliferation. Five variable kinds of breast cancer cells and one normal breast cell MCF-10A were treated with different doses of isomangiferin for 48 hours. Data are shown as mean±standard error of the mean (SEM) from triplicate experiments. The cell viability was determined by MTS assay according to Methods. (B) Isomangiferin supressed MDA-MB-231 migration. MDA-MB-231 cells were seeded in transwell chambers and then deprived of fetal bovine serum (FBS) for 6 hours. After that, cells were treated with 10 ng/mL vascular endothelial growth factor (VEGF) and indicated concentrations of isomangiferin. The migrated cells were quantified by manual counting by using an microscope (scale bar, 10 µm). Values are shown as mean±SEM of three independent experiments. (C) Isomangiferin imparied MDA-MB-231 invasion. After MDA-MB-231 cells seeded in the upper chamber of a transwell which has been pre-coated with Matrigel, the cells were deprived of FBS and then treated with different concentrations of isomangiferin. After 8 to 10 hours, the MDA-MB-231 cells that invaded through the membrane from the upper chamber to the bottom chamber were quantified when stimulated by the bottom chamber which was added VEGF (scale bar, 20 µm). Values are shown as mean±SEM of three independent experiments. *p<0.05 vs. MCF-10A; †p<0.01 vs. MCF-10A; ‡p<0.05 vs. 0 µM isomangiferin; §p<0.01 vs. 0 µM isomangiferin; ∥p<0.001 vs. 0 µM isomangiferin.
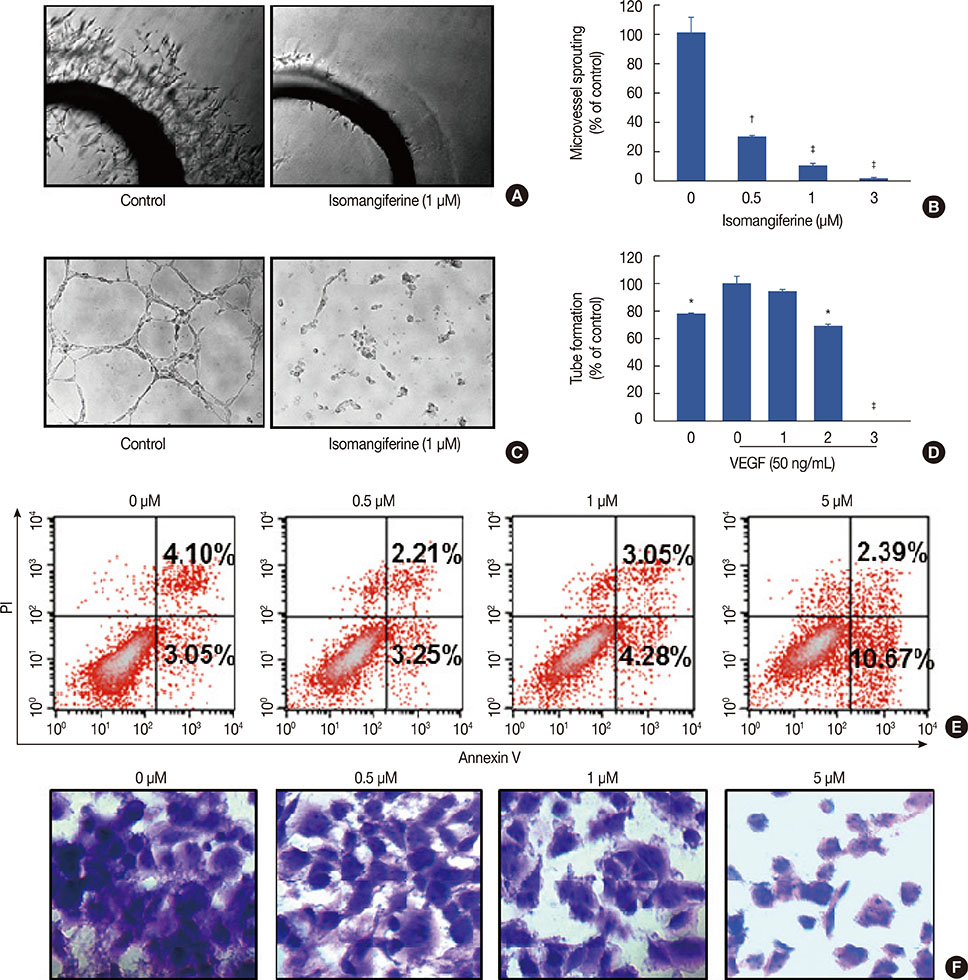
Figure 3 Isomangiferin inhibited the vascular endothelial growth factor (VEGF)-induced microvessel sprouting and tube formation of human umbilical vein endothelial cells (HUVECs) and induced MDA-MB-231 breast cancer cells apoptosis and adhension. (A) From Sprague-Dawley rats, the aortic segments were isolated and then were placed in growth factor-reduced Matrigel-covered wells. To stimulate angiogenesis, the vessel rings were treated with 50 ng/mL VEGF in the presence or absence of isomangiferin for 5 days. Representative photographs of endothelial cell sprouts forming branching cords from the margins of aortic rings under the treatment of 1 µM isomangiferin. (B) Sprouts were scored from 0 (least positive) to 5 (most positive) in a double-blinded manner. Values are shown as mean±standard error of the mean (SEM) of three independent experiments. (C) Isomangiferin inhibited the VEGF-induced tube formation of HUVEC tubes. HUVECs were placed in 96-well plates coated with Matrigel (6.5×103 per well). After 10 hours, cells were fixed, and tubular structures were photographed (magnification, ×100). (D) The statistic results of Figure 3C. Values are shown as mean±SEM of three independent experiments. (E) After incubation with different doses of isomangiferin for 36 hours, MDA-MB-231 cells were collected and apoptosis was assessed by annexin V-fluorescein isothiocyanate and prodidium iodide (annexin V/PI) staining and flow cytometry (n=3). (F) MDA-MB-231 cells were incubated with indicated doses of isomangiferin for 1 hour. Following incubation, cells were exposed and adhered to Matrigel. After 1% crystal violet stained, the number and phenotype of adhering cells was determined by microscope (magnification, ×400). *p<0.05 vs. control; †p<0.01 vs. control; ‡p<0.001 vs. control.
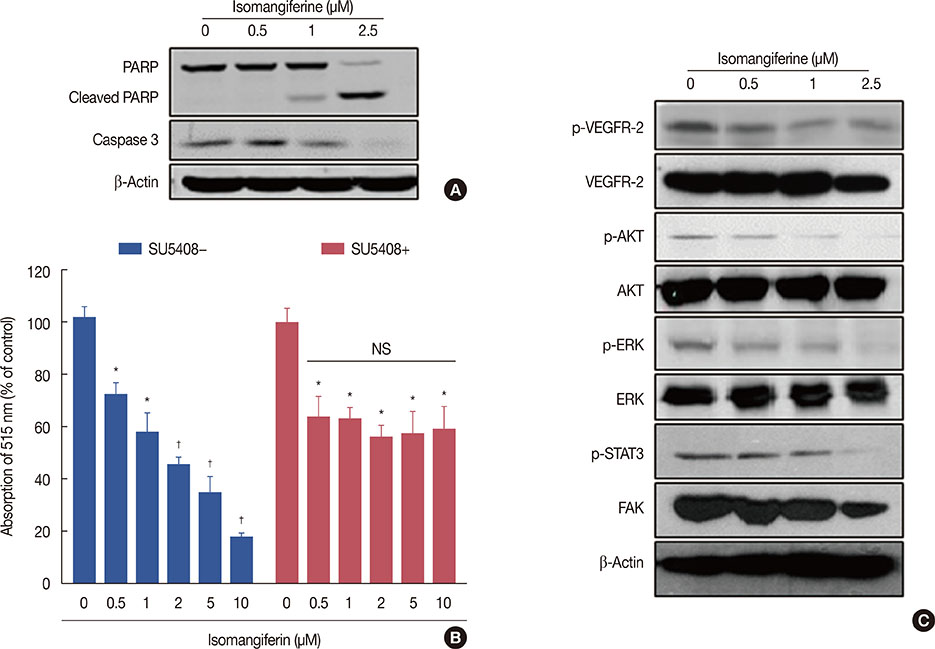
Figure 4 Isomangiferin suppresses breast cancer through inhibiting the vascular endothelial growth factor receptor 2 (VEGFR-2) signaling pathway. (A) Isomangiferin induced cell apoptosis. Proteins from MDA-MB-231 cells treated with indicated concentrations of isomangiferin for 24 hours were submitted to Western blot for the immunoblotting of caspase-3 and cleaved poly ADP-ribose polymerase (cleaved PARP). (B) Isomangiferin's ihibiton on breast cancer cell proliferation was dependent on VEGFR-2 activity. 100 nM SU5408 was used or not to block VEGFR-2's activity and then MDA-MB-231 cells were treated with isomangiferin. The cell proliferation was assessed by MTS assay. Values are shown as mean±standard error of the mean of three independent experiments. (C) Isomangiferin suppressed the activation of VEGFR-2 in human umbilical vein endothelial cells (HUVECs). The activation of VEGFR-2 was analyzed by Western blot and probed with indicated antibodies. Western blot was conducted in the way that described in Methods and specific antibodies were used accordingly. PARP=poly ADP-ribose polymerase; NS=not significant; p-AKT=phosphorylated protein kinase B, p-PKB/p-AKT; AKT=protein kinase B, PKB/AKT; p-ERK=phosphorylated extracellular regulated protein kinases; ERK=extracellular regulated protein kinases; p-STAT3=phosphorylated signal transducer and activator of transcription 3; FAK=focal adhesion kinase. *p<0.05 vs. control; †p<0.01 vs. control.
Reference
-
1. Jandial R, Hoshide R, Waters JD, Somlo G. Operative and therapeutic advancements in breast cancer metastases to the brain. Clin Breast Cancer. Epub 2017 Oct 7. DOI: 10.1016/j.clbc.2017.10.002.
Article2. Moreno AC, Lin YH, Bedrosian I, Shen Y, Stauder MC, Smith BD, et al. Use of regional nodal irradiation and its association with survival for women with high-risk, early stage breast cancer: a national cancer database analysis. Adv Radiat Oncol. 2017; 2:291–300.
Article3. Santoni M, Guerra F, Conti A, Lucarelli A, Rinaldi S, Belvederesi L, et al. Incidence and risk of cardiotoxicity in cancer patients treated with targeted therapies. Cancer Treat Rev. 2017; 59:123–131.
Article4. Zhou J, Wang H, Zhang H, Lutz AM, Tian L, Hristov D, et al. VEGFR2-targeted three-dimensional ultrasound imaging can predict responses to antiangiogenic therapy in preclinical models of colon cancer. Cancer Res. 2016; 76:4081–4089.
Article5. Pan Y, Zheng M, Zhong L, Yang J, Zhou S, Qin Y, et al. A preclinical evaluation of SKLB261, a multikinase inhibitor of EGFR/Src/VEGFR2, as a therapeutic agent against pancreatic cancer. Mol Cancer Ther. 2015; 14:407–418.
Article6. Maione P, Sgambato A, Casaluce F, Sacco PC, Santabarbara G, Rossi A, et al. The role of the antiangiogenetic ramucirumab in the treatment of advanced non small cell lung cancer. Curr Med Chem. 2017; 24:3–13.
Article7. Longatto-Filho A, Pinheiro C, Martinho O, Moreira MA, Ribeiro LF, Queiroz GS, et al. Molecular characterization of EGFR, PDGFRA and VEGFR2 in cervical adenosquamous carcinoma. BMC Cancer. 2009; 9:212.
Article8. Dunna NR, Kandula V, Girdhar A, Pudutha A, Hussain T, Bandaru S, et al. High affinity pharmacological profiling of dual inhibitors targeting RET and VEGFR2 in inhibition of kinase and angiogeneis events in medullary thyroid carcinoma. Asian Pac J Cancer Prev. 2015; 16:7089–7095.
Article9. Szabo E, Schneider H, Seystahl K, Rushing EJ, Herting F, Weidner KM, et al. Autocrine VEGFR1 and VEGFR2 signaling promotes survival in human glioblastoma models in vitro and in vivo. Neuro Oncol. 2016; 18:1242–1252.
Article10. Sato H, Takeda Y. VEGFR2 expression and relationship between tumor neovascularization and histologic characteristics in oral squamous cell carcinoma. J Oral Sci. 2009; 51:551–557.
Article11. Zhang J, Liu C, Shi W, Yang L, Zhang Q, Cui J, et al. The novel VEGF receptor 2 inhibitor YLL545 inhibits angiogenesis and growth in breast cancer. Oncotarget. 2016; 7:41067–41080.
Article12. Zhu X, Zhou W. The emerging regulation of VEGFR-2 in triple-negative breast cancer. Front Endocrinol (Lausanne). 2015; 6:159.
Article13. Yan JD, Liu Y, Zhang ZY, Liu GY, Xu JH, Liu LY, et al. Expression and prognostic significance of VEGFR-2 in breast cancer. Pathol Res Pract. 2015; 211:539–543.
Article14. Islam MN, Yoo HH, Lee J, Nam JW, Seo EK, Jin C, et al. Simultaneous determination of bioactive xanthone glycosides and norlignans from ethanolic extract of Anemarrhena asphodeloides by liquid chromatography. J AOAC Int. 2008; 91:1271–1277.
Article15. Satish Rao BS, Sreedevi MV, Nageshwar Rao B. Cytoprotective and antigenotoxic potential of Mangiferin, a glucosylxanthone against cadmium chloride induced toxicity in HepG2 cells. Food Chem Toxicol. 2009; 47:592–600.
Article16. Shen J, Song G, An M, Li X, Wu N, Ruan K, et al. The use of hollow mesoporous silica nanospheres to encapsulate bortezomib and improve efficacy for non-small cell lung cancer therapy. Biomaterials. 2014; 35:316–326.
Article17. Song G, Shen J, Jiang F, Hu R, Li W, An L, et al. Hydrophilic molybdenum oxide nanomaterials with controlled morphology and strong plasmonic absorption for photothermal ablation of cancer cells. ACS Appl Mater Interfaces. 2014; 6:3915–3922.
Article18. Li C, Hu J, Li W, Song G, Shen J. Combined bortezomib-based chemotherapy and p53 gene therapy using hollow mesoporous silica nanospheres for p53 mutant non-small cell lung cancer treatment. Biomater Sci. 2016; 5:77–88.
Article19. Shen J, Ma H, Zhang T, Liu H, Yu L, Li G, et al. Magnolol inhibits the growth of non-small cell lung cancer via inhibiting microtubule polymerization. Cell Physiol Biochem. 2017; 42:1789–1801.
Article20. Shen J, Sheng X, Chang Z, Wu Q, Wang S, Xuan Z, et al. Iron metabolism regulates p53 signaling through direct heme-p53 interaction and modulation of p53 localization, stability, and function. Cell Rep. 2014; 7:180–193.
Article21. Shen J, Zhang S, Li Y, Zhang W, Chen J, Zhang M, et al. p14(ARF) inhibits the functions of adenovirus E1A oncoprotein. Biochem J. 2011; 434:275–285.
Article22. Yamada KH, Kang H, Malik AB. Antiangiogenic therapeutic potential of peptides derived from the molecular motor KIF13B that transports VEGFR2 to plasmalemma in endothelial cells. Am J Pathol. 2017; 187:214–224.
Article23. Varga A, Gyulavári P, Greff Z, Futosi K, Németh T, Simon-Szabó L, et al. Targeting vascular endothelial growth factor receptor 2 and protein kinase D1 related pathways by a multiple kinase inhibitor in angiogenesis and inflammation related processes in vitro. PLoS One. 2015; 10:e0124234.
Article24. Chen H, Cong Q, Du Z, Liao W, Zhang L, Yao Y, et al. Sulfated fucoidan FP08S2 inhibits lung cancer cell growth in vivo by disrupting angiogenesis via targeting VEGFR2/VEGF and blocking VEGFR2/Erk/VEGF signaling. Cancer Lett. 2016; 382:44–52.
Article25. DeWire M, Fouladi M, Turner DC, Wetmore C, Hawkins C, Jacobs C, et al. An open-label, two-stage, phase II study of bevacizumab and lapatinib in children with recurrent or refractory ependymoma: a collaborative ependymoma research network study (CERN). J Neurooncol. 2015; 123:85–91.
Article26. Liu H, Wu B, Pan G, He L, Li Z, Fan M, et al. Metabolism and pharmacokinetics of mangiferin in conventional rats, pseudo-germ-free rats, and streptozotocin-induced diabetic rats. Drug Metab Dispos. 2012; 40:2109–2118.
Article27. Zhao J, Zhang B, Li S, Zeng L, Chen Y, Fang J. Mangiferin increases Nrf2 protein stability by inhibiting its ubiquitination and degradation in human HL60 myeloid leukemia cells. Int J Mol Med. 2014; 33:1348–1354.
Article28. Dilshara MG, Kang CH, Choi YH, Kim GY. Mangiferin inhibits tumor necrosis factor-alpha-induced matrix metalloproteinase-9 expression and cellular invasion by suppressing nuclear factor-kappaB activity. BMB Rep. 2015; 48:559–564.
Article29. Núñez Selles AJ, Daglia M, Rastrelli L. The potential role of mangiferin in cancer treatment through its immunomodulatory, anti-angiogenic, apoptopic, and gene regulatory effects. Biofactors. 2016; 42:475–491.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Expression of Vascular Endothelial Growth Factor-C in Breast Carcinoma
- The Relationship between PTEN Tumor Suppressor Gene and Vascular Endothelial Growth Factor-Mediated Angiogenesis in Breast Cancer
- Pivotal role of vascular endothelial growth factor pathway in tumor angiogenesis
- Zerumbone, Sesquiterpene Photochemical from Ginger, Inhibits Angiogenesis
- Thrombospondin-1 and Inhibition of Tumor Growth
