Induction Mechanism of PD-L1 (Programmed Cell Death-ligand 1) in Sepsis
- Affiliations
-
- 1Division of Pulmonary and Critical Care Medicine, Department of Internal Medicine and Lung Institute, Medical Research Center, Seoul National University College of Medicine, Seoul, Korea. sangmin2@snu.ac.kr
Abstract
- PD-L1 is expressed in a variety of antigen-presenting cells and provides T cell tolerance via ligation with its receptor PD-1 and B7-1 on T cells. Stimulation with lipopolysaccharide (LPS) can increase the level of PD-L1 expression in B cells and macrophages, which suggests that this molecule plays a role in the immunosuppression observed in severe sepsis. The aim of this study was to identify which of the downstream pathways of TLR4 are involved in the up-regulation of PD-L1 by LPS in macrophages. Flow cytometry was used to examine the expression of PD-L1 in RAW 264.7 macrophages stimulated with LPS. The following chemical inhibitors were used to evaluate the role of each pathway: LY294002 for PI3K/Akt, SB202190 for p38 MAPK, and U0126 for MEK. LPS induced the expression of PD-L1 in a time- and dose-dependent manner. Transfection of siRNA for TLR4 suppressed the induction of PD-L1. Pretreatment with LY294002 and SB202190 decreased the level of PD-L1 expression but U0126 did not. Overall, the PI3K/Akt and p38 MAPK pathways are involved in the up-regulation of PD-L1 expression in RAW 264.7 macrophages stimulated with LPS.
Keyword
- PD-L1; LPS; Sepsis; Macrophage
MeSH Terms
-
Antigen-Presenting Cells
B-Lymphocytes
Butadienes
Chromones
Flow Cytometry
Imidazoles
Immunosuppression
Ligation
Macrophages
Morpholines
Nitriles
p38 Mitogen-Activated Protein Kinases
Pyridines
RNA, Small Interfering
Sepsis
T-Lymphocytes
Transfection
Up-Regulation
Butadienes
Chromones
Imidazoles
Morpholines
Nitriles
Pyridines
RNA, Small Interfering
p38 Mitogen-Activated Protein Kinases
Figure
-
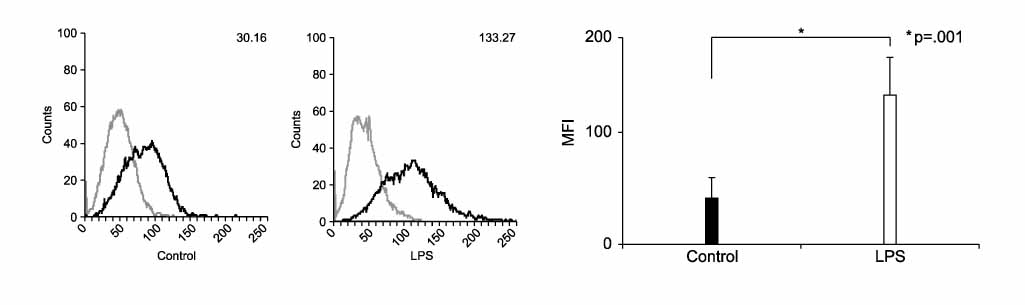
Figure 1 PD-L1 is up-regulated by LPS in mouse macrophage cell line. RAW 264.7 cells were treated with LPS (1.0 µg/ml) for 24 hrs. Surface expressions of PD-L1 were measured by flow cytometry and expressed via MFI (mean fluorescence intensity). Number: mean fluorscence intensity (MFI). Gray: isotype control, Black: PD-L1.
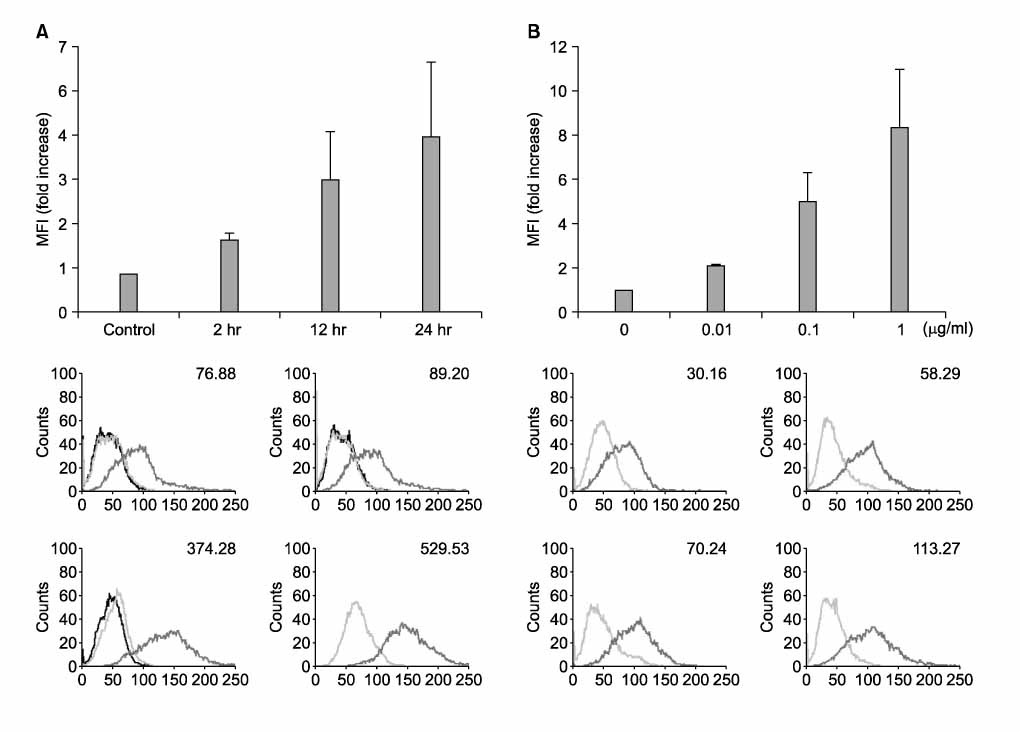
Figure 2 PD-L1 is up-regulated by LPS in time-dependent & concentration-dependent manner. RAW 264.7 cells were treated with LPS (1.0 µg/ml) for the indicated times (A) and with the indicated concentrations of LPS for 24 hrs (B). Surface expressions of PD-L1 were measured by flow cytometry and expressed via MFI.
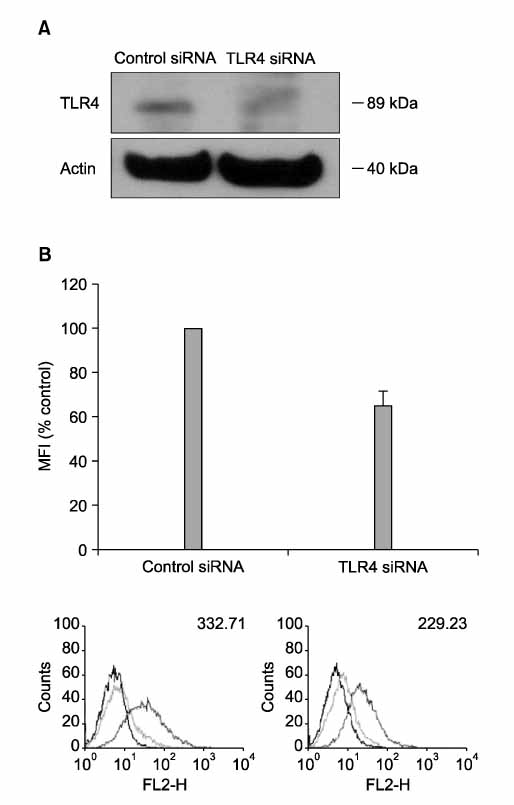
Figure 3 Blocking of TLR4 with siRNA suppress LPS-induced PD-L1 expression. RAW 264.7 cells were transfected with control siRNA and siRNA for TLR4. The levels of TLR4 in cellular extracts were detected by Western blot analysis (A). After cells were treated with LPS (1.0 µg/ml) for 24 hrs, surface expressions of PD-L1 were measured by flow cytometry and expressed via MFI (B).
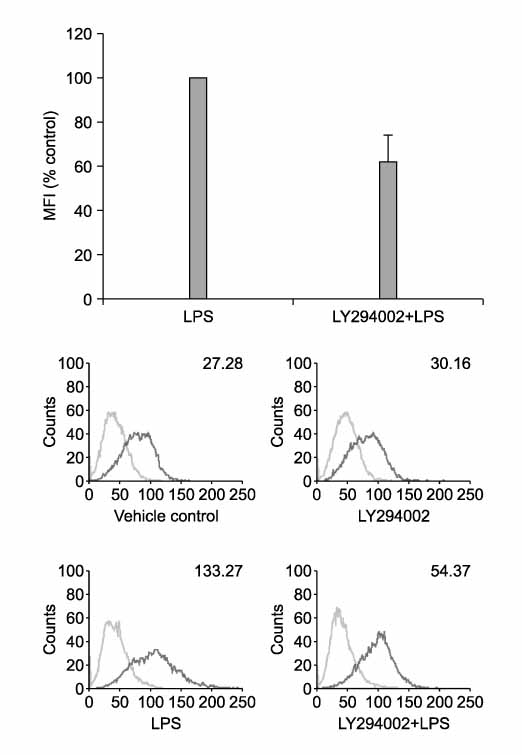
Figure 4 PI3K/Akt pathway is involved in up-regulation of PD-L1 by LPS. RAW 264.7 cells were pretreated with LY294002 (50 µM) for 2 hrs and then stimulated with LPS for 24 hrs. Surface expressions of PD-L1 were measured by flow cytometry and expressed via MFI. Cell viability by MTT assay: > 90% of vehicle control.
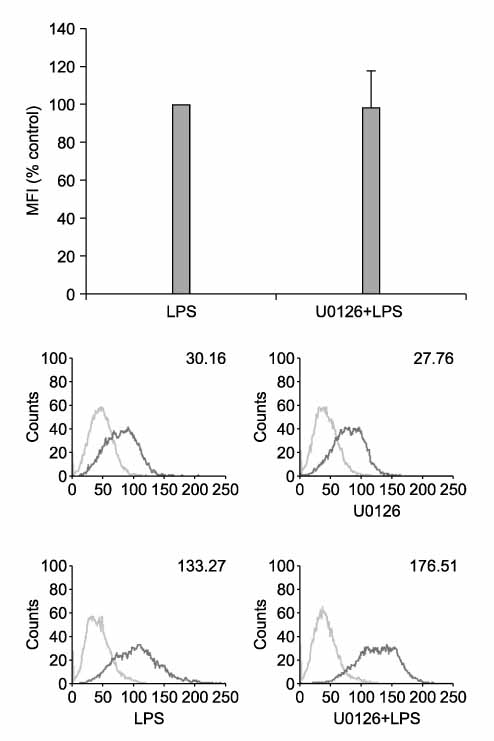
Figure 5 MEK/ERK pathway has no role in up-regulation of PD-L1 by LPS. RAW 264.7 cells were pretreated with U0126 (10 µM) for 2 hrs and then stimulated with LPS for 24 hrs. Surface expressions of PD-L1 were measured by flow cytometry and expressed via MFI. Cell viability by MTT assay: > 90% of vehicle control.
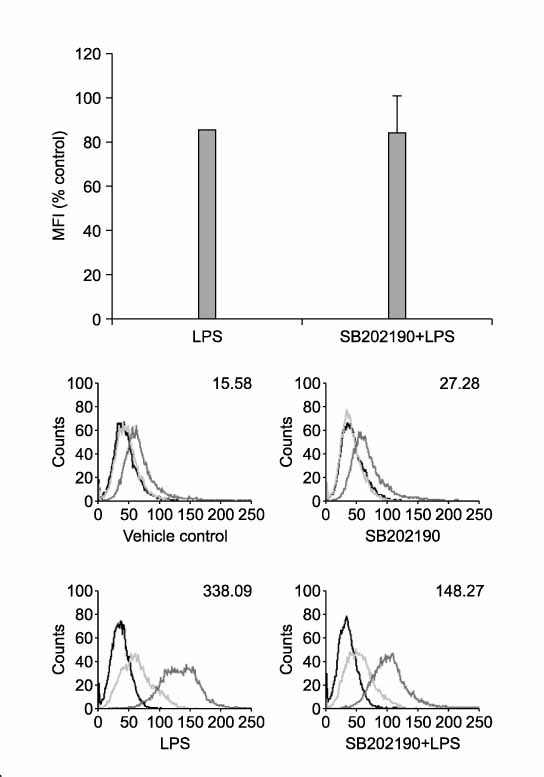
Figure 6 p38 MAPK pathway is involved in up-regulation of PD-L1 by LPS. RAW 264.7 cells were pretreated with SB202190 (10 µM) for 2 hrs and then stimulated with LPS for 24 hrs. Surface expressions of PD-L1 were measured by flow cytometry and expressed via MFI. Cell viability by MTT assay: > 90% of vehicle control.
Reference
-
1. Angus DC, Linde-Zwirble WT, Lidicker J, Clermont G, Carcillo J, Pinsky MR. Epidemiology of severe sepsis in the United States: analysis of incidence, outcome, and associated costs of care. Crit Care Med. 2001. 29:1303–1310.2. Bone RC, Fisher CJ Jr, Clemmer TP, Slotman GJ, Metz CA, Balk RA. A controlled clinical trial of high-dose methylprednisolone in the treatment of severe sepsis and septic shock. N Engl J Med. 1987. 317:653–658.3. Bone RC. Sir Isaac Newton, sepsis, SIRS, and CARS. Crit Care Med. 1996. 24:1125–1128.4. Hotchkiss RS, Karl IE. The pathophysiology and treatment of sepsis. N Engl J Med. 2003. 348:138–150.5. Okazaki T, Honjo T. The PD-1-PD-L pathway in immunological tolerance. Trends Immunol. 2006. 27:195–201.6. Iwai Y, Terawaki S, Ikegawa M, Okazaki T, Honjo T. PD-1 inhibits antiviral immunity at the effector phase in the liver. J Exp Med. 2003. 198:39–50.7. Liang SC, Latchman YE, Buhlmann JE, Tomczak MF, Horwitz BH, Freeman GJ, et al. Regulation of PD-1, PD-L1, and PD-L2 expression during normal and autoimmune responses. Eur J Immunol. 2003. 33:2706–2716.8. Wiendl H, Mitsdoerffer M, Schneider D, Chen L, Lochmuller H, Melms A, et al. Human muscle cells express a B7-related molecule, B7-H1, with strong negative immune regulatory potential: a novel mechanism of counterbalancing the immune attack in idiopathic inflammatory myopathies. FASEB J. 2003. 17:1892–1894.9. Sharpe AH, Wherry EJ, Ahmed R, Freeman GJ. The function of programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nat Immunol. 2007. 8:239–245.10. Nishimura H, Minato N, Nakano T, Honjo T. Immunological studies on PD-1 deficient mice: implication of PD-1 as a negative regulator for B cell responses. Int Immunol. 1998. 10:1563–1572.11. Nishimura H, Nose M, Hiai H, Minato N, Honjo T. Development of lupus-like autoimmune diseases by disruption of the PD-1 gene encoding an ITIM motif-carrying immunoreceptor. Immunity. 1999. 11:141–151.12. Ansari MJ, Salama AD, Chitnis T, Smith RN, Yagita H, Akiba H, et al. The programmed death-1 (PD-1) pathway regulates autoimmune diabetes in nonobese diabetic (NOD) mice. J Exp Med. 2003. 198:63–69.13. Guleria I, Khosroshahi A, Ansari MJ, Habicht A, Azuma M, Yagita H, et al. A critical role for the programmed death ligand 1 in fetomaternal tolerance. J Exp Med. 2005. 202:231–237.14. Thompson RH, Gillett MD, Cheville JC, Lohse CM, Dong H, Webster WS, et al. Costimulatory B7-H1 in renal cell carcinoma patients: indicator of tumor aggressiveness and potential therapeutic target. Proc Natl Acad Sci U S A. 2004. 101:17174–17179.15. Ohigashi Y, Sho M, Yamada Y, Tsurui Y, Hamada K, Ikeda N, et al. Clinical significance of programmed death-1 ligand-1 and programmed death-1 ligand-2 expression in human esophageal cancer. Clin Cancer Res. 2005. 11:2947–2953.16. Yamazaki T, Akiba H, Iwai H, Matsuda H, Aoki M, Tanno Y, et al. Expression of programmed death 1 ligands by murine T cells and APC. J Immunol. 2002. 169:5538–5545.17. Loke P, Allison JP. PD-L1 and PD-L2 are differentially regulated by Th1 and Th2 cells. Proc Natl Acad Sci U S A. 2003. 100:5336–5341.18. Annane D, Bellissant E, Cavaillon JM. Septic shock. Lancet. 2005. 365:63–78.19. Lee SK, Seo SH, Kim BS, Kim CD, Lee JH, Kang JS, et al. IFN-gamma regulates the expression of B7-H1 in dermal fibroblast cells. J Dermatol Sci. 2005. 40:95–103.20. Keir ME, Latchman YE, Freeman GJ, Sharpe AH. Programmed death-1 (PD-1):PD-ligand 1 interactions inhibit TCR-mediated positive selection of thymocytes. J Immunol. 2005. 175:7372–7379.21. Saunders PA, Hendrycks VR, Lidinsky WA, Woods ML. PD-L2:PD-1 involvement in T cell proliferation, cytokine production, and integrin-mediated adhesion. Eur J Immunol. 2005. 35:3561–3569.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Evaluation of circulating PD-1 and PD-L1 as diagnostic biomarkers in dogs with tumors
- PD-L1 Testing in Non-small Cell Lung Cancer: Past, Present, and Future
- Immunohistochemical expression of programmed death-ligand 1 and CD8 in glioblastomas
- Programmed cell death-ligand 1 assessment in urothelial carcinoma: prospect and limitation
- PD-L1 as a Biomarker in Gastric Cancer Immunotherapy
