Bis is Induced by Oxidative Stress via Activation of HSF1
- Affiliations
-
- 1Department of Biochemistry, College of Medicine, The Catholic University of Korea, Seoul 137-701, Korea. leejh@catholic.ac.kr
- 2Cancer Research Institute, College of Medicine, The Catholic University of Korea, Seoul 137-701, Korea.
- 3Cancer Evolution Research Center, College of Medicine, The Catholic University of Korea, Seoul 137-701, Korea.
- KMID: 2285539
- DOI: http://doi.org/10.4196/kjpp.2014.18.5.403
Abstract
- The Bis protein is known to be involved in a variety of cellular processes including apoptosis, migration, autophagy as well as protein quality control. Bis expression is induced in response to a number of types of stress, such as heat shock or a proteasome inhibitor via the activation of heat shock factor (HSF)1. We report herein that Bis expression is increased at the transcriptional level in HK-2 kidney tubular cells and A172 glioma cells by exposure to oxidative stress such as H2O2 treatment and oxygen-glucose deprivation, respectively. The pretreatment of HK-2 cells with N-acetyl cysteine, suppressed Bis induction. Furthermore, HSF1 silencing attenuated Bis expression that was induced by H2O2, accompaniedby increase in reactive oxygen species (ROS) accumulation. Using a series of deletion constructs of the bis gene promoter, two putative heat shock elements located in the proximal region of the bis gene promoter were found to be essential for the constitutive expression is as well as the inducible expression of Bis. Taken together, our results indicate that oxidative stress induces Bis expression at the transcriptional levels via activation of HSF1, which might confer an expansion of antioxidant capacity against pro-oxidant milieu. However, the possible role of the other cis-element in the induction of Bis remains to be determined.
Keyword
MeSH Terms
Figure
-
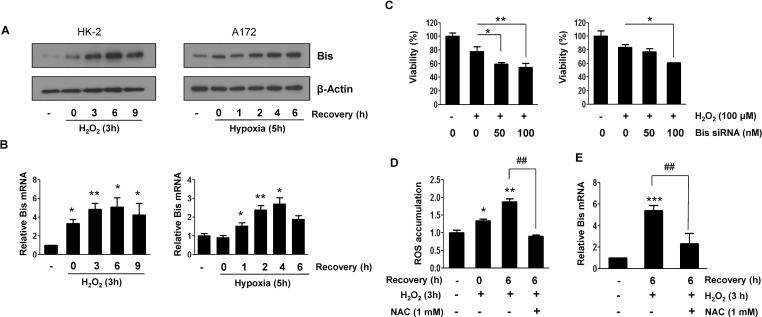
Fig. 1 Induction of Bis expression by oxidative stress in HK-2 and A172 cells. (A) Western blot assay for Bis expression in HK-2 (left column) and A172 (right column) cells after exposure to H2O2 and OGD, respectively, and then incubated in the normal medium for indicated times. Beta-actin expression was used as a loading control. (B) The relative levels of Bis mRNA from HK-2 (left column) or A172 (right column) cells at the indicated conditions were determined by quantitative real time RT-PCR analysis after normalizing with beta-actin mRNA level in the same sample. The mean values from three independent experiments are shown with SD. The value from the control cells before exposure to oxidative stress is designated as 1.0. *p<0.05 and **p<0.01 vs. the value from control cells. (C) HK-2 (left column) or A172 (right column) cells were transfected with control or Bis specific siRNA with indicated doses for 48 h and then treated with 100 µM of H2O2 for 3 h followed by additional 6 h in normal medium. The relative cell viability was determined using MTT assay as described in METHODS section. Values from triplicate experiments are provided as mean±SE. *p<0.05 and **p<0.01 vs. the value from control cells treated only with H2O2. (D) ROS accumulation was determined in HK-2 cells after treatment of H2O2 by measuring DCF-DA fluorescence intensity using flow cytometric analysis. 1 mM of NAC was pretreated before exposure to H2O2. Fold changes in the mean from three experiments are provided as mean±SE. *p<0.05 and **p<0.01 vs. the value from control cells, ##p<0.01 vs. the value in the absence of NAC. (E) Effect of NAC on the induction of Bis mRNA was determined by pretreatment of NAC prior to H2O2 treatment and then Bis mRNA level was determined as in (B). The mean values from four independent experiments are present with SD. ***p<0.001 vs. the value from control cells, ##p<0.01 vs. the value in the absence of NAC.
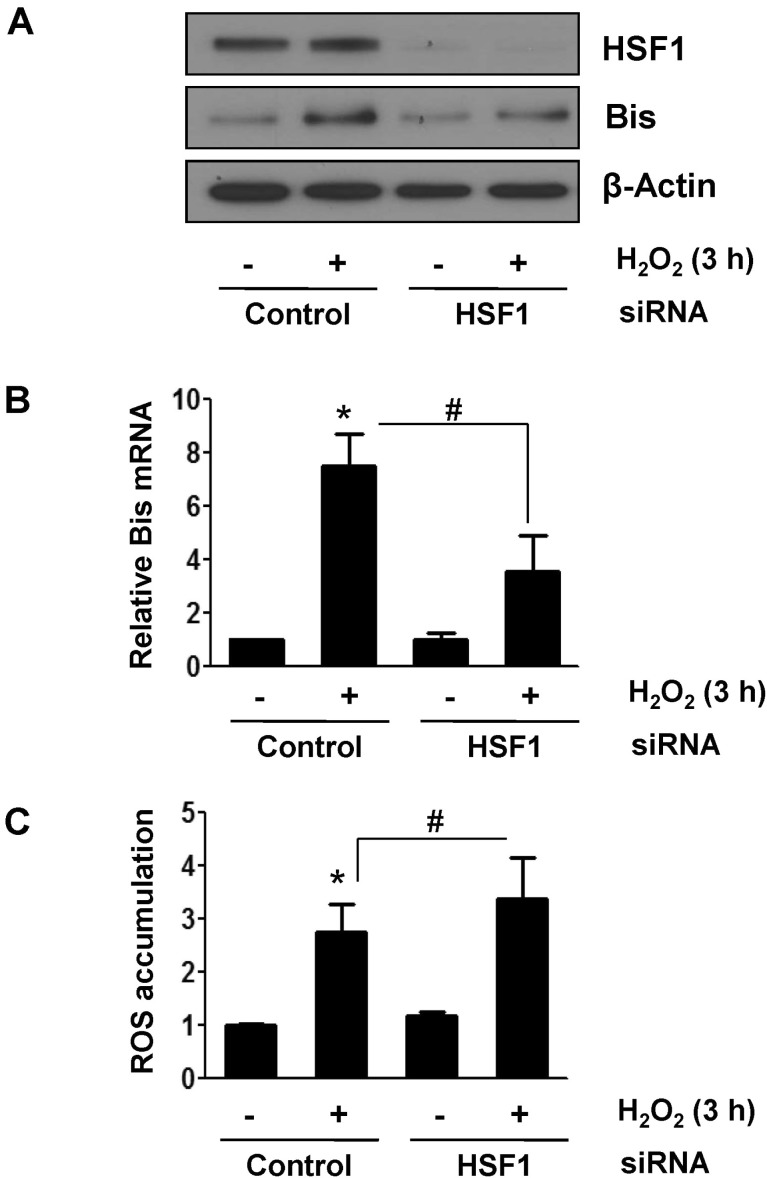
Fig. 2 HSF1 suppressed induction of Bis mRNA upon oxidative stress. HK-2 cells were transfected with control or HSF1-specific siRNA and incubated with H2O2 for 3 h followed by additional 6 h in normal medium. HSF1 and Bis expressions were evaluated by Western assay (A) and by real-time RT-PCR (B) as in Fig. 1. (C) Effect of downregulation of HSF1 on ROS accumulation upon H2O2 treatment was examined by DCF-DA staining and FACS analysis. Data are presented with fold changes in the mean intensities from three independent experiments with SE. *p<0.05 vs. the value from control cells, #p<0.05 vs. the value from the control siRNA-treated cells.
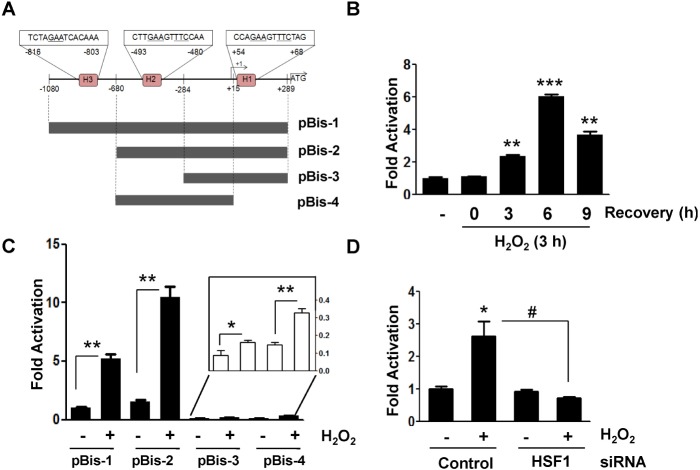
Fig. 3 Identification of oxidative stress-responsive region in the Bis promoter (A) Schematic diagram of the deletion mutants for proximal region of Bis promoter, which were cloned into pGL3 basic vector.The relative position to the transcription site (+1) and the locations of putative HSE (H) are shown. (B) Transcriptional activation of pBis-1 was determined at the indicated time points following treatment of H2O2 after normalization with renilla activity. Luciferase activity before exposure to H2O2 is designated at 1.0. The mean value from triplicate experiments are present with SD. **p<0.01 and ***p<0.001 vs. the value from control cells. (C) Fold activation of various deletion mutants of Bis promoter in response to H2O2 treatment was shown. The luciferase activityof pBis-1 before exposure to H2O2 is designated at 1.0. The values for pBis-3 and pBis-4 were provided as a magnified graph (inlet). The mean values from triplicate experiments are present with SD. *p<0.05 and **p<0.01 vs. the value from H2O2-untreated cells in each construct. (D) Effect of HSF1 knockdown on the expression of pBis-2 was shown. Expression of HSF1 was suppressed by transfection of HSF1 specific siRNA for 24 h and then HK-2 cells were transfected with pBis-2 construct as described in Materials and Methods section. Fold change in luciferase activity compared with that of control HK-2 cells are presented as mean value with SD. *p<0.05 vs. the value from H2O2-untreated control cells, #p<0.05 vs. the value from the control siRNA-treated cells.
Reference
-
1. Lee JH, Takahashi T, Yasuhara N, Inazawa J, Kamada S, Tsujimoto Y. Bis, a Bcl-2-binding protein that synergizes with Bcl-2 in preventing cell death. Oncogene. 1999; 18:6183–6190. PMID: 10597216.
Article2. Takayama S, Xie Z, Reed JC. An evolutionarily conserved family of Hsp70/Hsc70 molecular chaperone regulators. J Biol Chem. 1999; 274:781–786. PMID: 9873016.
Article3. Doong H, Price J, Kim YS, Gasbarre C, Probst J, Liotta LA, Blanchette J, Rizzo K, Kohn E. CAIR-1/BAG-3 forms an EGF-regulated ternary complex with phospholipase C-gamma and Hsp70/Hsc70. Oncogene. 2000; 19:4385–4395. PMID: 10980614.4. Doong H, Rizzo K, Fang S, Kulpa V, Weissman AM, Kohn EC. CAIR-1/BAG-3 abrogates heat shock protein-70 chaperone complexmediated protein degradation: accumulation of poly-ubiquitinated Hsp90 client proteins. J Biol Chem. 2003; 278:28490–28500. PMID: 12750378.5. Kassis JN, Guancial EA, Doong H, Virador V, Kohn EC. CAIR-1/BAG-3 modulates cell adhesion and migration by downregulating activity of focal adhesion proteins. Exp Cell Res. 2006; 312:2962–2971. PMID: 16859681.
Article6. Suzuki M, Iwasaki M, Sugio A, Hishiya A, Tanaka R, Endo T, Takayama S, Saito T. BAG3 (BCL2-associated athanogene 3) interacts with MMP-2 to positively regulate invasion by ovarian carcinoma cells. Cancer Lett. 2011; 303:65–71. PMID: 21316839.
Article7. Fontanella B, Birolo L, Infusini G, Cirulli C, Marzullo L, Pucci P, Turco MC, Tosco A. The co-chaperone BAG3 interacts with the cytosolic chaperonin CCT: new hints for actin folding. Int J Biochem Cell Biol. 2010; 42:641–650. PMID: 20018251.
Article8. Iwasaki M, Tanaka R, Hishiya A, Homma S, Reed JC, Takayama S. BAG3 directly associates with guanine nucleotide exchange factor of Rap1, PDZGEF2, and regulates cell adhesion. Biochem Biophys Res Commun. 2010; 400:413–418. PMID: 20800573.
Article9. Carra S, Seguin SJ, Lambert H, Landry J. HspB8 chaperone activity toward poly(Q)-containing proteins depends on its association with Bag3, a stimulator of macroautophagy. J Biol Chem. 2008; 283:1437–1444. PMID: 18006506.
Article10. Gamerdinger M, Hajieva P, Kaya AM, Wolfrum U, Hartl FU, Behl C. Protein quality control during aging involves recruitment of the macroautophagy pathway by BAG3. EMBO J. 2009; 28:889–901. PMID: 19229298.
Article11. Crippa V, Sau D, Rusmini P, Boncoraglio A, Onesto E, Bolzoni E, Galbiati M, Fontana E, Marino M, Carra S, Bendotti C, De Biasi S, Poletti A. The small heat shock protein B8 (HspB8) promotes autophagic removal of misfolded proteins involved in amyotrophic lateral sclerosis (ALS). Hum Mol Genet. 2010; 19:3440–3456. PMID: 20570967.
Article12. Seo YJ, Jeon MH, Lee JH, Lee YJ, Youn HJ, Ko JH, Lee JH. Bis induces growth inhibition and differentiation of HL-60 cells via up-regulation of p27. Exp Mol Med. 2005; 37:624–630. PMID: 16391524.
Article13. Yoon JS, Lee MY, Lee JS, Park CS, Youn HJ, Lee JH. Bis is involved in glial differentiation of p19 cells induced by retinoic Acid. Korean J Physiol Pharmacol. 2009; 13:251–256. PMID: 19885044.
Article14. De Marco M, Turco MC, Rosati A. BAG3 protein is induced during cardiomyoblast differentiation and modulates myogenin expression. Cell Cycle. 2011; 10:850–852. PMID: 21311226.
Article15. Zhu H, Liu P, Li J. BAG3: a new therapeutic target of human cancers? Histol Histopathol. 2012; 27:257–261. PMID: 22237703.16. Pagliuca MG, Lerose R, Cigliano S, Leone A. Regulation by heavy metals and temperature of the human BAG-3 gene, a modulator of Hsp70 activity. FEBS Lett. 2003; 541:11–15. PMID: 12706811.
Article17. Chen L, Wu W, Dentchev T, Zeng Y, Wang J, Tsui I, Tobias JW, Bennett J, Baldwin D, Dunaief JL. Light damage induced changes in mouse retinal gene expression. Exp Eye Res. 2004; 79:239–247. PMID: 15325571.
Article18. Tabuchi Y, Ando H, Takasaki I, Feril LB Jr, Zhao QL, Ogawa R, Kudo N, Tachibana K, Kondo T. Identification of genes responsive to low intensity pulsed ultrasound in a human leukemia cell line Molt-4. Cancer Lett. 2007; 246:149–156. PMID: 16678341.
Article19. Wang HQ, Liu HM, Zhang HY, Guan Y, Du ZX. Transcriptional upregulation of BAG3 upon proteasome inhibition. Biochem Biophys Res Commun. 2008; 365:381–385. PMID: 17996194.
Article20. Jacobs AT, Marnett LJ. HSF1-mediated BAG3 expression attenuates apoptosis in 4-hydroxynonenal-treated colon cancer cells via stabilization of anti-apoptotic Bcl-2 proteins. J Biol Chem. 2009; 284:9176–9183. PMID: 19179333.
Article21. Song S, Kole S, Precht P, Pazin MJ, Bernier M. Activation of heat shock factor 1 plays a role in pyrrolidine dithiocarbamatemediated expression of the co-chaperone BAG3. Int J Biochem Cell Biol. 2010; 42:1856–1863. PMID: 20692357.
Article22. Franceschelli S, Rosati A, Lerose R, De Nicola S, Turco MC, Pascale M. Bag3 gene expression is regulated by heat shock factor 1. J Cell Physiol. 2008; 215:575–577. PMID: 18286539.
Article23. Du ZX, Zhang HY, Meng X, Gao YY, Zou RL, Liu BQ, Guan Y, Wang HQ. Proteasome inhibitor MG132 induces BAG3 expression through activation of heat shock factor 1. J Cell Physiol. 2009; 218:631–637. PMID: 19006120.
Article24. Gentilella A, Passiatore G, Deshmane S, Turco MC, Khalili K. Activation of BAG3 by Egr-1 in response to FGF-2 in neuroblastoma cells. Oncogene. 2008; 27:5011–5018. PMID: 18469860.
Article25. Li C, Li S, Kong DH, Meng X, Zong ZH, Liu BQ, Guan Y, Du ZX, Wang HQ. BAG3 is upregulated by c-Jun and stabilizes JunD. Biochim Biophys Acta. 2013; 1833:3346–3354. PMID: 24140207.
Article26. Gentilella A, Khalili K. Autoregulation of co-chaperone BAG3 gene transcription. J Cell Biochem. 2009; 108:1117–1124. PMID: 19777443.
Article27. Lee MY, Kim SY, Shin SL, Choi YS, Lee JH, Tsujimoto Y, Lee JH. Reactive astrocytes express bis, a bcl-2-binding protein, after transient forebrain ischemia. Exp Neurol. 2002; 175:338–346. PMID: 12061864.
Article28. Cho KO, Lee KE, Youn DY, Jeong KH, Kim JY, Yoon HH, Lee JH, Kim SY. Decreased vulnerability of hippocampal neurons after neonatal hypoxia-ischemia in bis-deficient mice. Glia. 2012; 60:1915–1929. PMID: 22907804.29. Jung SE, Kim YK, Youn DY, Lim MH, Ko JH, Ahn YS, Lee JH. Down-modulation of Bis sensitizes cell death in C6 glioma cells induced by oxygen-glucose deprivation. Brain Res. 2010; 1349:1–10. PMID: 20599823.
Article30. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001; 25:402–408. PMID: 11846609.31. Wu C. Heat shock transcription factors: structure and regulation. Annu Rev Cell Dev Biol. 1995; 11:441–469. PMID: 8689565.
Article32. Lim JH, Youn DY, Yoo HJ, Yoon HH, Kim MY, Chung S, Kim YS, Chang YS, Park CW, Lee JH. Aggravation of diabetic nephropathy in BCL-2 interacting cell death suppressor (BIS)-haploinsufficient mice together with impaired induction of superoxide dismutase (SOD) activity. Diabetologia. 2014; 57:214–223. PMID: 24078136.
Article33. Anckar J, Sistonen L. Regulation of HSF1 function in the heat stress response: implications in aging and disease. Annu Rev Biochem. 2011; 80:1089–1115. PMID: 21417720.
Article34. Gidalevitz T, Prahlad V, Morimoto RI. The stress of protein misfolding: from single cells to multicellular organisms. Cold Spring Harb Perspect Biol. 2011; 3:pii: a009704.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- ERK-mediated phosphorylation of BIS regulates nuclear translocation of HSF1 under oxidative stress
- Effect of BIS depletion on HSF1-dependent transcriptional activation in A549 non-small cell lung cancer cells
- Epilepsy and Oxidative Stress
- Roles of the complement system in alcohol-induced liver disease
- Oxidative stress and endometriosis
