Effects of Wnt-1 on the Growth and Apoptosis of FRTL-5 Cells
- Affiliations
-
- 1Department of Internal Medicine, Asan Medical Center, University of Ulsan College of Medicine, Korea.
- 2Asan Institute for Life Sciences, University of Ulsan College of Medicine, Korea.
- KMID: 2178240
- DOI: http://doi.org/10.3803/jkes.2007.22.1.35
Abstract
-
BACKGROUND: Wnt proteins are major signaling molecules involved in embryonic induction, generation of cell polarity and the cell fate decision. A central player in the Wnt signaling pathways is beta-catenin. Several studies have suggested that the Wnt/beta-catenin signaling pathway may be involved in the physiologic/pathologic control of thyroid cell growth and function.
METHODS
We investigated the effect of thyroid-stimulating hormone (TSH) on the expression of Wnt proteins in FRTL-5 cells. To evaluate the effect of Wnt-1 on FRTL-5 cells growth, we isolated a stable cell line that overexpressed Wnt-1 (W1), and a vector-transfected cell clone (V3) was used as a control. We investigated the differences in the cellular growth rate, the cell cycle and cell apoptosis in the W1 and V3 cell lines.
RESULTS
TSH caused a significant increase in the Wnt-1 level and a pronounced decrease in both the active and total beta-catenin levels in the FRTL-5 cells. The growth rate, the percentage of cells in the S/G2/M phase and the c-myc level were significantly higher in the W1 cells compared with the V3 cells. There was no change in the beta-catenin level and the cyclin D1 level in the W1 cells compared with the V3 cells. The cellular apoptosis induced by actinomycin-D seemed to be significantly decreased because the level of bcl-2 was increased in the W1 cells compared with the V3 cells.
CONCLUSION
The FRTL-5 cells expressed Wnt-1 protein, and TSH increased the Wnt-1 expression, and it paradoxically decreased beta-catenin in the FRTL-5 cells. Overexpression of Wnt-1 in the FRTL-5 cells increased cell growth and it decreased apoptosis. Growth stimulation by Wnt-1 overexpression was not mediated by beta-catenin (the canonical Wnt pathway), but seemed to be mediated by activation of the Wnt/Ca2+ pathway, which involves an increased c-myc level. Suppression of apoptosis with Wnt-1 overexpression was due to the increased bcl-2 level.
Keyword
MeSH Terms
Figure
-
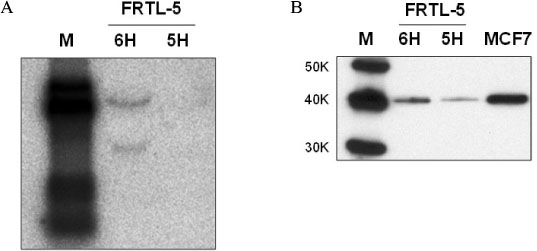
Fig. 1 Wnt-1 expression in FRTL-5 cells being increased by TSH. FRTL-5 cells were grown in the absence (5H media, '5H') or presence (6H media, '6H') of TSH for 5 days. A, Total cellular RNA was subjected to Northern blot analyses using Wnt-1 probe. B, 20 µg of cell lysates were subjected to SDS-PAGE and Western blot analysis using Wnt-1 antibody. MCF7, a breast cancer cell line known to over-produce Wnt-1 protein, was used as positive control.
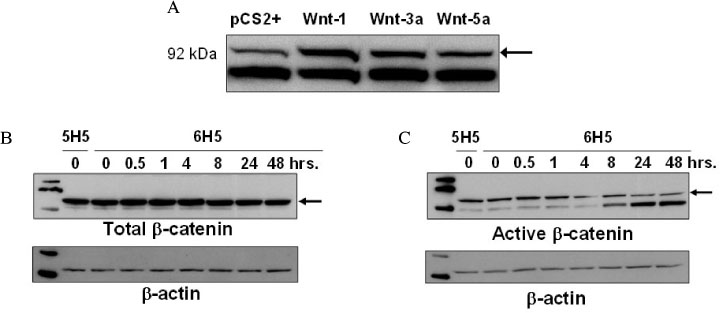
Fig. 2 Total and active beta-catenin level in FRTL-5 cells being affected by TSH. A, Lysates of FRTL-5 cells transfected with Wnt-1, Wnt-3a or Wnt-5a expression vectors were subjected to SDS-PAGE and Western blot analysis using antibody against 'active beta-catenin'. B, Lysates of FRTL-5 cells grown in the absence (5H media, '5H') or presence (6H media, '6H') of TSH were subjected to SDS-PAGE and Western blot analysis using antibody against total beta-catenin. C, Lysates of FRTL-5 cells grown in the absence (5H media, '5H') or presence (6H media, '6H') of TSH were subjected to SDS-PAGE and Western blot analysis using antibody against active beta-catenin. Arrows indicate target protein (total or active beta-catenin).

Fig. 3 The growth rate of FRTL-5 cells stably transfected with Wnt-1. Growth rates of FRTL-5 cells stably transfected with Wnt-1 expression vector (W1) or control vector (V3) were compared at indicated time points using MTT assay. Background absorbance (cell culture medium only) was subtracted to obtain actual absorbance. Absorbance values were plotted relative to baseline absorbance (i.e., absorbance at day 0). All values are mean ± standard deviation.
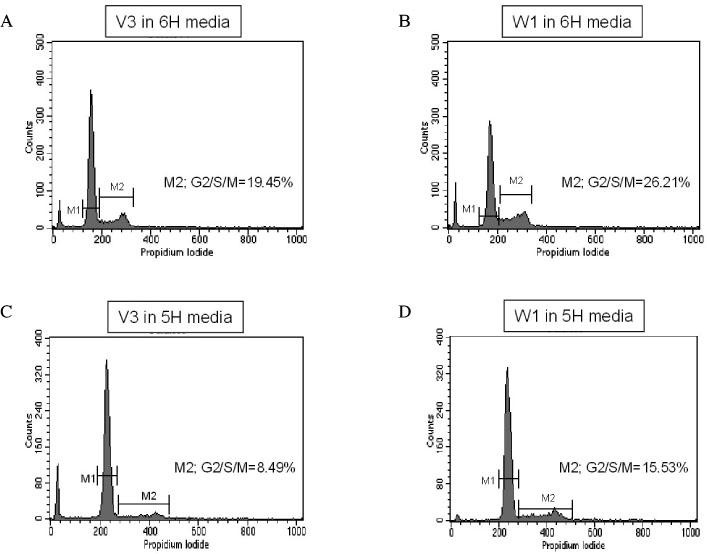
Fig. 4 Cell cycle analysis in stable transfectant overexpressing Wnt-1. Cell cycle analysis of Wnt-1 overexpressing cells (W1) and vector-transfected cells (V3) was performed using flow cytometry. DNA content of cells stained with propidium iodide was measured and analyzed. Cell cycle distribution of V3 clone grown in 6H media (A), maintained in 5H media for 6 days (B), W1 clone grown in 6H media (C), and maintained in 5H media for 6 days (D) are shown. M1 indicates cells in G1 phase and M2 indicates cells in G2/S/M phase.
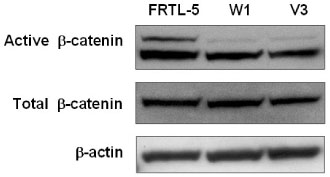
Fig. 5 Beta-catenin level in Wnt-1 transfected FRTL-5 cells. 20 µg of cell lysates of Wnt-1 transfected (W1) or vector-transfected (V3) FRTL-5 cells and FRTL-5 cells grown in the presence of TSH (6H media) were subjected to SDS-PAGE and Western blot analysis using antibodies against 'active beta-catenin' and 'total beta-catenin'.
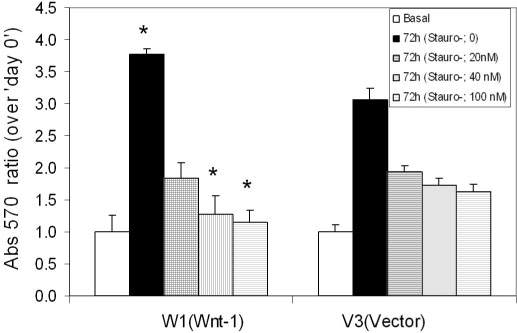
Fig. 6 Effect of staurosporin, a PKC inhibitor, on the growth rate of FRTL-5 cells stably trasfected with Wnt-1 or control vector. Growth rates of FRTL-5 cells stably transfected with Wnt-1 expression vector (W1) or control vector (V3) were compared in the presence of staurosporin or control (DMSO) using MTT assay. Background absorbance (cell culture medium only) was subtracted to obtain actual absorbance. Absorbance values at 72 hours were plotted relative to baseline absorbance (i.e., absorbance at day 0). All values are mean ± standard deviation of 6 independent wells. * indicates P< 0.05 compared to V3 cells treated with the same concentration of staurosporin.
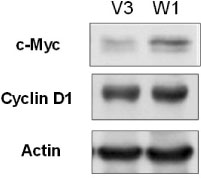
Fig. 7 Protein expressions of c-myc, cyclin D, and actin of Wnt-1 transfected (W1) and vector-transfected (V3) FRTL-5 cells. Expressions of c-myc, cyclin D1, and actin of W1 and V3 were measured by Western blot analysis.
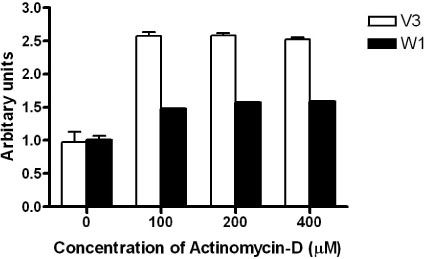
Fig. 8 Effects of actinomycin-D on apoptosis of Wnt-1 transfected (W1) and vector-transfected (V3) FRTL-5 cells. Apoptosis was measured by Cell death detection ELISA plus kit (Roche Diagnostics). W1 and V3 were treated by 0, 200, and 400 µM of actinomycin-D for 48 hours, respectively.
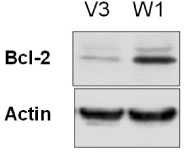
Fig. 9 Protein expressions of bcl-2 and actin of Wnt-1 transfected (W1) and vector-transfected (V3) FRTL-5 cells. Expressions of bcl-2 and actin of W1 and V3 were measured by Western blot analysis.
Reference
-
1. Cadigan KM, Nusse R. Wnt signaling: a common theme in animal development. Genes Dev. 1997. 11:3286–3305.2. Moon RT, Kohn AD, De Ferrari GV, Kaykas A. WNT and beta-catenin signalling: diseases and therapies. Nat Rev Genet. 2004. 5:691–701.3. Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science. 2004. 303:1483–1487.4. Jamora C, Fuchs E. Intercellular adhesion, signalling and the cytoskeleton. Nat Cell Biol. 2002. 4:E101–E108.5. Gumbiner BM. Regulation of cadherin adhesive activity. J Cell Biol. 2000. 148:399–404.6. Miller JR, Hocking AM, Brown JD, Moon RT. Mechanism and function of signal transduction by the Wnt/beta-catenin and Wnt/Ca2+ pathways. Oncogene. 1999. 18:7860–7872.7. Polakis P. Wnt signaling and cancer. Genes Dev. 2000. 14:1837–1851.8. Polakis P. Casein kinase 1: a Wnt'er of disconnect. Curr Biol. 2002. 12:R499–R501.9. Morin PJ. Beta-catenin signaling and cancer. Bioessays. 1999. 21:1021–1030.10. Webster MT, Rozycka M, Sara E, Davis E, Smalley M, Young N, Dale TC, Wooster R. Sequence variants of the axin gene in breast, colon, and other cancers: an analysis of mutations that interfere with GSK3 binding. Genes Chromosomes Cancer. 2000. 28:443–453.11. Clevers H. Axin and hepatocellular carcinomas. Nat Genet. 2000. 24:206–208.12. Polakis P. The adenomatous polyposis coli (APC) tumor suppressor. Biochim Biophys Acta. 1997. 1332:F127–F147.13. Helmbrecht K, Kispert A, von WR, Brabant G. Identification of a Wnt/beta-catenin signaling pathway in human thyroid cells. Endocrinology. 2001. 142:5261–5266.14. Cerrato A, Fulciniti F, Avallone A, Benincasa G, Palombini L, Grieco M. Beta- and gamma-catenin expression in thyroid carcinomas. J Pathol. 1998. 185:267–272.15. Huang SH, Wu JC, Chang KJ, Liaw KY, Wang SM. Expression of the cadherin-catenin complex in well-differentiated human thyroid neoplastic tissue. Thyroid. 1999. 9:1095–1103.16. Garcia-Rostan G, Tallini G, Herrero A, D'Aquila TG, Carcangiu ML, Rimm DL. Frequent mutation and nuclear localization of beta-catenin in anaplastic thyroid carcinoma. Cancer Res. 1999. 59:1811–1815.17. Garcia-Rostan G, Camp RL, Herrero A, Carcangiu ML, Rimm DL, Tallini G. Beta-catenin dysregulation in thyroid neoplasms: down-regulation, aberrant nuclear expression, and CTNNB1 exon 3 mutations are markers for aggressive tumor phenotypes and poor prognosis. Am J Pathol. 2001. 158:987–996.18. Kohn LD, Saji M, Akamizu T, Ikuyama S, Isozaki O, Kohn AD, Santisteban P, Chan JY, Bellur S, Rotella CM. Receptors of the thyroid: the thyrotropin receptor is only the first violinist of a symphony orchestra. Adv Exp Med Biol. 1989. 261:151–209.19. Bidey SP, Lambert A, Robertson WR. Thyroid cell growth, differentiation and function in the FRTL-5 cell line: a survey. J Endocrinol. 1988. 119:365–376.20. Kikkawa F, Gonzalez FJ, Kimura S. Characterization of a thyroid-specific enhancer located 5.5 kilobase pairs upstream of the human thyroid peroxidase gene. Mol Cell Biol. 1990. 10:6216–6224.21. Shimura H, Okajima F, Ikuyama S, Shimura Y, Kimura S, Saji M, Kohn LD. Thyroid-specific expression and cyclic adenosine 3',5'-monophosphate autoregulation of the thyrotropin receptor gene involves thyroid transcription factor-1. Mol Endocrinol. 1994. 8:1049–1069.22. Yost C, Torres M, Miller JR, Huang E, Kimelman D, Moon RT. The axis-inducing activity, stability, and subcellular distribution of beta-catenin is regulated in Xenopus embryos by glycogen synthase kinase 3. Genes Dev. 1996. 10:1443–1454.23. Yang-Snyder J, Miller JR, Brown JD, Lai CJ, Moon RT. A frizzled homolog functions in a vertebrate Wnt signaling pathway. Curr Biol. 1996. 6:1302–1306.24. McMahon AP, Moon RT. Ectopic expression of the proto-oncogene int-1 in Xenopus embryos leads to duplication of the embryonic axis. Cell. 1989. 58:1075–1084.25. Emami KH, Nguyen C, Ma H, Kim DH, Jeong KW, Eguchi M, Moon RT, Teo JL, Kim HY, Moon SH, Ha JR, Kahn M. A small molecule inhibitor of beta-catenin/CREB-binding protein transcription [corrected]. Proc Natl Acad Sci U S A. 2004. 101:12682–12687.26. Suzuki K, Mori A, Ishii KJ, Saito J, Singer DS, Klinman DM, Krause PR, Kohn LD. Activation of target-tissue immune-recognition molecules by double-stranded polynucleotides. Proc Natl Acad Sci U S A. 1999. 96:2285–2290.27. Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods. 1983. 65:55–63.28. van NM, van de WM, Clevers H. Identification of two novel regulated serines in the N terminus of beta-catenin. Exp Cell Res. 2002. 276:264–272.29. Lu Z, Hunter T. Wnt-independent beta-catenin transactivation in tumor development. Cell Cycle. 2004. 3:571–573.30. Tetsu O, McCormick F. Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature. 1999. 398:422–426.31. Shtutman M, Zhurinsky J, Simcha I, Albanese C, D'Amico M, Pestell R, Ben-Ze'ev A. The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl Acad Sci U S A. 1999. 96:5522–5527.32. He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT, Morin PJ, Vogelstein B, Kinzler KW. Identification of c-MYC as a target of the APC pathway. Science. 1998. 281:1509–1512.33. Pelengaris S, Khan M, Evan G. c-MYC: more than just a matter of life and death. Nat Rev Cancer. 2002. 2:764–776.34. Isozaki O, Kohn LD. Control of c-fos and c-myc proto-oncogene induction in rat thyroid cells in culture. Mol Endocrinol. 1987. 1:839–848.35. Sheldahl LC, Park M, Malbon CC, Moon RT. Protein kinase C is differentially stimulated by Wnt and Frizzled homologs in a G-protein-dependent manner. Curr Biol. 1999. 9:695–698.36. Slusarski DC, Corces VG, Moon RT. Interaction of Wnt and a Frizzled homologue triggers G-protein-linked phosphatidylinositol signalling. Nature. 1997. 390:410–413.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Effect of Estrogen on H2O2 Induced Apoptosis of FRTL-5 Cells
- Effect of ceramide on apoptosis and phospholipase D activity in FRTL-5 thyroid cells
- Pseudolaric acid B inhibits PAX2 expression through Wnt signaling and induces BAX expression, therefore promoting apoptosis in HeLa cervical cancer cells
- Overexpression of USF Increases TGF-beta1 Protein Levels, But G1 Phase Arrest was not Induced in FRTL-5 Cells
- Effect of Lithium on Na+/I- Symporter Gene Expression in Rat Thyroid FRTL-5 Cells
