House Dust Mite-Derived Chitin Enhances Th2 Cell Response to Inhaled Allergens, Mainly via a TNF-α-Dependent Pathway
- Affiliations
-
- 1Institute of Convergence Medicine, Ewha Womans University School of Medicine and Medical Center, Seoul, Korea. juinea@ewha.ac.kr
- 2Department of Internal Medicine, Gacheon University College of Medicine, Incheon, Korea.
- 3Department of Life Sciences, Pohang University of Science and Technology (POSTECH), Pohang, Korea.
- 4Academy of Immunology and Microbiology (AIM), Institute for Basic Science (IBS), Pohang, Korea.
- 5Department of Internal Medicine, Dankook University College of Medicine, Cheonan, Korea.
- KMID: 2165920
- DOI: http://doi.org/10.4168/aair.2016.8.4.362
Abstract
- PURPOSE
Chitin is a potent adjuvant in the development of immune response to inhaled allergens in the airways. According to other studies, chitin is known as multi-faced adjuvants which can induce Th2 responses. Recently, we found that TNF-α is a key mediator in the development of Th2 cell response to inhaled allergens. Here, we evaluated the immunologic mechanisms in the development of airway hypersensitivity to inhaled allergens, enhanced by house dust mite (HDM)-derived chitin.
METHODS
The role of TNF-α and TLRs was evaluated in an airway hypersensitivity mouse model induced by a sensitization with an allergen (ovalbumin, OVA) and HDM-derived chitin using mice with the null mutation of target genes.
RESULTS
The present study showed that airway sensitization with HDM-derived chitin plus OVA enhanced OVA-induced airway inflammation v. OVA alone. This phenotype was associated with the increased expression of Th1, Th2, and Th17 cytokines and also with the enhanced production of OVA-specific IgE, IgG1, and IgG2a. As for T cell responses, OVA-specific Th2 cell response, enhanced by chitin, was abolished by the treatment of chitinase, whereas Th1 and Th17 cell responses enhanced by this treatment. Moreover, the null mutation of the TNF-α gene revealed similar effects as the chitinase treatment. In contrast, all the OVA-specific T cell responses, enhanced by chitin, were blocked by the absence of TLR2, but not of TLR1, TLR4, or TLR6.
CONCLUSIONS
In conclusion, these data suggest that HDM-derived chitin may enhance airway hypersensitivity to inhaled allergens, via the TLR2-dependent pathway, and that chitin-induced TNF-α can be a key mediator in the development of Th2 cell response to inhaled allergens.
Keyword
MeSH Terms
Figure
-
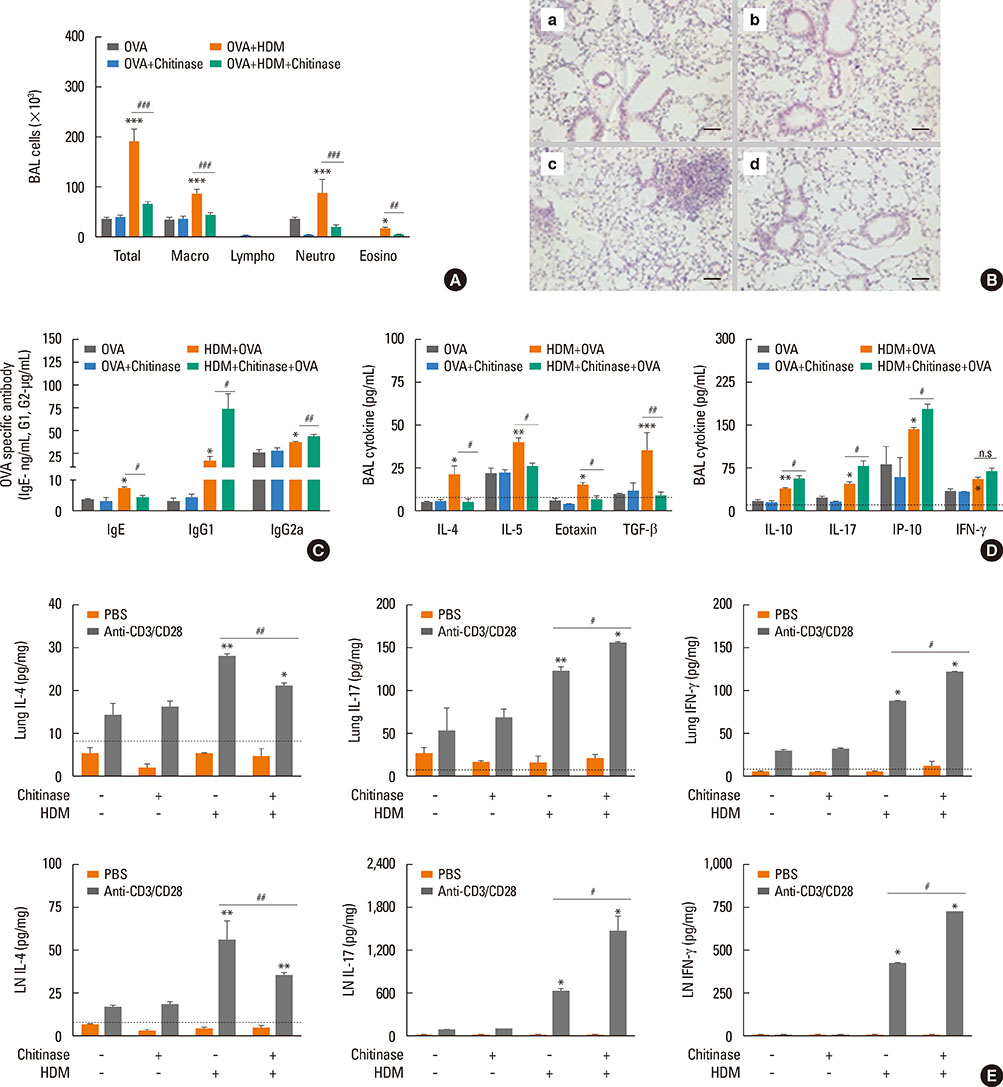
Fig. 1 Th2-dominant airway inflammation induced by HDM extracts plus OVA is down-regulated by chitinase treatment. For all panels, wild-type mice (C57BL/6 background) were sensitized with 75 µg of OVA+100 µg of HDM with or without chitinase, then challenged with OVA (50 µg) alone. The evaluations were performed 24 hours after last OVA challenge. *P<0.05; **P<0.01; ***P<0.001 relative to individual control groups, OVA or OVA+Chitinase, respectively; #P<0.05; ##P<0.01; ###P<0.001; n.s, not significant. Data are presented as the mean±SEM (n=5 mice per group) are from a single experiment representative of at least 3 independent experiments. (A) Bronchoalveolar lavage (BAL) cellularity after acute allergen challenge. (B) Lung histologic findings (a, OVA; b, OVA+chitinase; c, OVA+HDM; d, OVA+HDM+chitinase; magnification: 100×, scale bar: 200 µm). (C) The production of OVA-specific IgE, IgG1, and IgG2a in serum after acute OVA challenge. (D) Cytokine productions of IL-4, IL-5, eotaxin, TGF-β, IL-10, IL-17, IP-10, and IFN-γ in BAL fluid after last allergen challenge. For (E) panel, cells were isolated from lung-draining lymph nodes (LN) and lung tissues, and incubated with PBS or anti-CD3 and CD28 antibodies for 12 hours. Levels of each cytokine were evaluated in supernatant fraction. (E) Levels of IL-4, IL-10, IL-17, and IFN-γ from lung cells, and lung-draining LN cells.
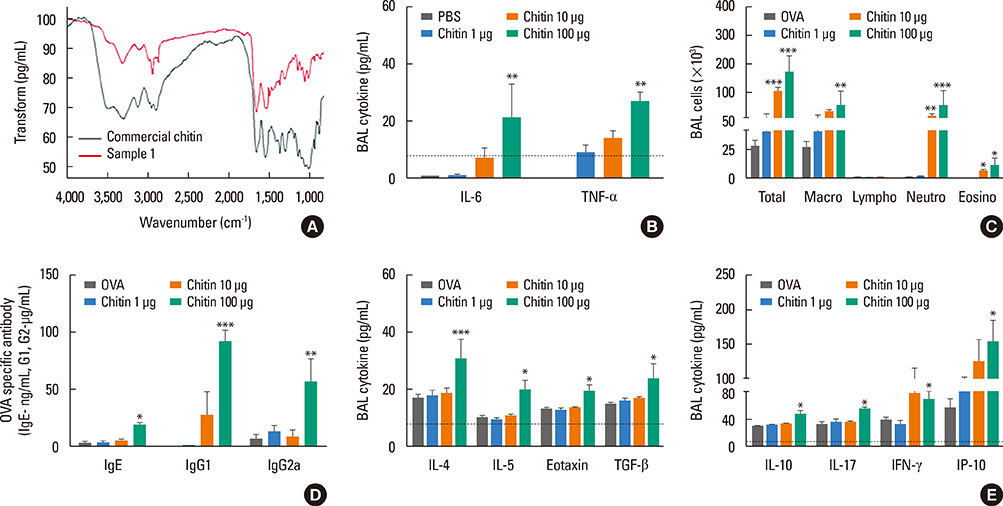
Fig. 2 Chitins extracted from HDM have an adjuvant effect to enhance adaptive immune response to inhaled allergens. (A) Fourier transform infrared (FTIR) spectroscopy of commercial crab chitin (commercial chitin) and HDM derived chitin (sample1). For (B) panel, HDM-derived chitin was sensitized to wild type (WT) mouse airways and then evaluated after 12 hours. For (C-E) panels, wild-type mice (C57BL/6 background) were sensitized with 75 µg of OVA, or 75 µg of OVA+1, 10, or 100 µg of HDM-derived chitin. Data are presented as the mean±SEM (n=5 mice per group) are from a single experiment representative of at least 2 independent experiments. *P<0.05; **P<0.01; ***P<0.001 relative to the OVA group; (B) Levels of IL-6, and TNF-α in BAL fluid after sensitization of each dose of HDM-derived chitin. For (C-E), mice were challenged with OVA (50 µg) alone and evaluated 24 hours after the last OVA challenge. (C) Bronchoalveolar lavage (BAL) cellularity after 24 hours of last OVA challenge. (D) Levels of OVA-specific IgE, IgG1, and IgG2a in serum after acute challenge. (E) Levels of IL-4, IL-5, eotaxin, TGF-β, IL-10, IL-17, IFN-γ, and IP-10 in BAL fluid after last acute OVA challenge.
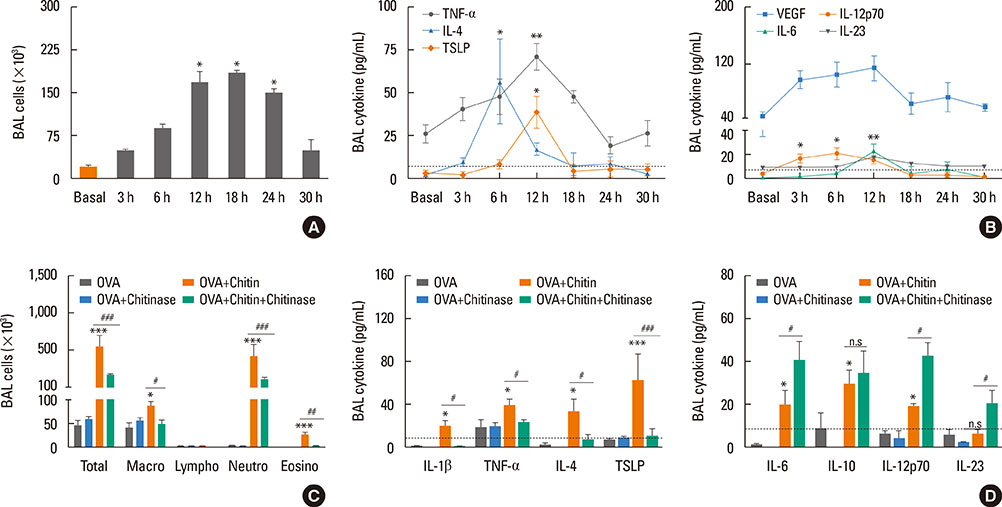
Fig. 3 Th2-type innate response, induced by HDM-derived chitin is abolished by chitinase treatment. Data are presented as the mean±SEM (n=5 mice per group) are from a single experiment representative of at least 3 independent experiments. *P<0.05; **P<0.01; ***P<0.001; n.s, not significant. relative to basal or the OVA group; For panel (A) and (B), 100 µg of HDM-derived chitin was sensitized once to wild type (WT) mouse airways and then evaluated at various time points. (A) Inflammatory cell infiltration of airway after HDM-derived chitin sensitization. (B) Levels of TNF-α, IL-4, TSLP, VEGF, IL-6, IL-12p70, and IL-23 in bronchoalveolar lavage (BAL) fluids. For (C-D), the evaluation was performed at 12 hours after HDM-derived chitin sensitization. (C) BAL cellularity after sensitization. (D) Levels of IL-1β, TNF-α, IL-4, TSLP, IL-6, IL-10, IL-12p70, and IL-23 in BAL fluid after sensitization.

Fig. 4 OVA-induced Th2-cell response enhanced by HDM-derived chitin is abolished by chitinase treatment. For all panels, wild-type (WT) mice (C57BL/6 background) were sensitized with 75 µg of OVA, 75 µg of OVA+chitinase, 75 µg of OVA+100 µg of HDM-derived chitin, or 75 µg of OVA+100 µg of HDM-derived chitin+chitinase, then challenged with OVA (50 µg) alone, and evaluated 24 hours after last OVA challenge. Data are presented as the mean±SEM (n=5 mice per group) are from a single experiment representative of at least 3 independent experiments. *P<0.05; **P<0.01; ***P<0.001 relative to the OVA group; #P<0.05; ##P<0.01; ###P<0.001. (A) Bronchoalveolar lavage (BAL) cellularity after acute OVA challenge. (B) Lung histologic findings (a, OVA; b, OVA+chitinase; c, OVA+HDM-derived chitin; d, OVA+HDM-derived chitin+chitinase; magnification: 100×, scale bar: 200 µm). (C) Levels of OVA-specific IgE, IgG1, IgG2a in serum after acute challenge. (D) Levels of IL-4, IL-5, eotaxin, TGF-β, IL-10, IL-17, IL-12p70, and IFN-γ in BAL fluid after acute challenge. For panels (E), cells were isolated from lung-draining lymph nodes (LNs) and lung tissues, and incubated with PBS or CD3 and CD28 antibodies for 12 hours. Levels of each cytokine were evaluated in supernatant fraction. (E) Levels of IL-4, IL-10, IL-17, and IFN-γ from lung and lung-draining LN cells.

Fig. 5 OVA-induced adaptive immune responses enhanced by HDM-derived chitin is abolished by the absence of TLR2. For all panels, TLR2-deficient mice and wild-type (WT) mice (C57BL/6 background) were sensitized with 75 µg of OVA, or 75 µg of OVA+100 µg of HDM derived chitin. *P<0.05; **P<0.01; ***P<0.001 relative to the wild-type OVA group; #P<0.05; ##P<0.01; ###P<0.001. Data are presented as the mean±SEM (n=5 mice per group) are from a single experiment representative of at least 2 independent experiments. For (A-C) panels, evaluations were performed after 12-hour sensitization. (A) Bronchoalveolar lavage (BAL) cellularity after sensitization. (B) Levels of IL-1β, TNF-α, IL-4, TSLP, IL-6, IL-10, IL-12p70, and IL-23 in BAL fluid after sensitization. For panels (C-H), mice were challenged with OVA (50 µg) alone, and evaluated 24 hours after last OVA challenge. (C) BAL cellularity after acute challenge. (D) Lung histologic findings (a, WT_OVA; b, TLR2 KO_OVA; c, WT_Chitin+OVA; d, TLR2 KO_OVA+chitin; magnification: 100×, scale bar: 200 µm). (E) Levels of OVA-specific IgE, IgG1, IgG2a in serum after acute challenge. (F) Levels of IL-4, IL-5, eotaxin, TGF-β, IL-10, IL-17, IFN-γ, and IP-10 in BAL fluid after acute challenge. For panels (G, H), cells were isolated from lung-draining lymph nodes (LNs) and lung tissues. (G) The levels of memory T-cell proliferation by stimulation of OVA in LN cells from each mouse. For (H), isolated cells were incubated with PBS or CD3 and CD28 antibodies for 12 hours. Levels of individual cytokines were evaluated in supernatant fraction. (H) Levels of IL-4, IL-10, IL-17, and IFN-γ from lung and lung-draining LN cells.
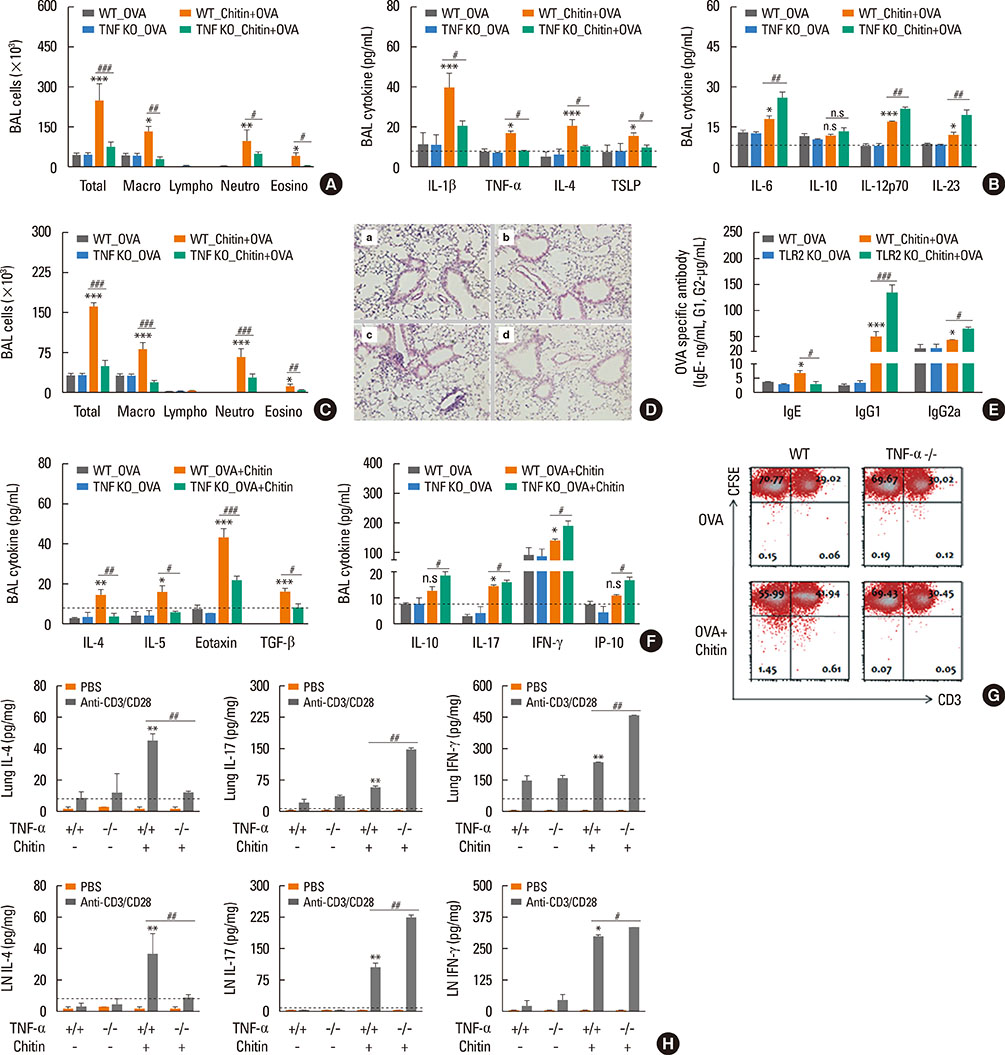
Fig. 6 OVA-induced Th2-cell response, enhanced by HDM-derived chitin is abolished by the absence of TNF-α. Th2 responses induced by airway sensitization with ovalbumin (OVA) plus HDM-derived chitin is diminished in the absence of TNF-α, whereas the Th1- and Th17-cell responses are enhanced. For all panels, TNF-α-deficient (TNF KO) mice and wild-type (WT) mice (C57BL/6 background) were sensitized with 75 µg of OVA, or 75 µg of OVA+100 µg of HDM-derived chitin. Data are presented as the mean±SEM (n=5 mice per group) are from a single experiment representative of at least two independent experiments. *P<0.05; **P<0.01; ***P<0.001 relative to the wild-type OVA group; #P<0.05; ##P<0.01; ###P<0.001; n.s, not significant. For panel (A) and (B), the evaluation was performed 12 hours after sensitization. (A) Bronchoalveolar lavage (BAL) cellularity after sensitization. (B) Levels of IL-1β, TNF-α, IL-4, TSLP, IL-6, IL-10, IL-12p70, and IL-23 in BAL fluid after sensitization. For (C-H), mice were challenged with OVA (50 µg) alone and evaluated 24 hours after last OVA challenge. (C) BAL cellularity after acute challenge. (D) Lung histologic findings (a, WT_OVA; b, TNF KO_OVA; c, WT_OVA+chitin; d, TNF KO_OVA+chitin; magnification: 100×, scale bar: 200 µm). (E) Levels of OVA-specific IgE, IgG1, and IgG2a in serum after acute challenge. (F) Levels of IL-4, IL-5, eotaxin, TGF-β, IL-10, IL-17, IP-10, and IFN-γ in BAL fluid after acute challenge. For panels (G, H), cells were isolated from lung-draining lymph nodes (LNs) and lung tissues. (G) The levels of memory T-cell proliferation by stimulation of OVA in LN cells from each mouse. For (H), isolated cells were incubated with PBS or CD3 and CD28 antibodies for 12 hours. Levels of individual cytokine were evaluated in supernatant fraction. (H) Levels of IL-4, IL-10, IL-17, and IFN-γ from lung and lung-draining LN cells.
Cited by 1 articles
-
Bcl11b Regulates IL-17 Through the TGF-β/Smad Pathway in HDM-Induced Asthma
Si Chen, Yuting Han, Hao Chen, Jing Wu, Min Zhang
Allergy Asthma Immunol Res. 2018;10(5):543-554. doi: 10.4168/aair.2018.10.5.543.
Reference
-
1. Dowse GK, Turner KJ, Stewart GA, Alpers MP, Woolcock AJ. The association between Dermatophagoides mites and the increasing prevalence of asthma in village communities within the Papua New Guinea highlands. J Allergy Clin Immunol. 1985; 75:75–83.2. Charpin D, Birnbaum J, Haddi E, Genard G, Lanteaume A, Toumi M, et al. Altitude and allergy to house-dust mites. A paradigm of the influence of environmental exposure on allergic sensitization. Am Rev Respir Dis. 1991; 143:983–986.3. Platts-Mills TA, Vervloet D, Thomas WR, Aalberse RC, Chapman MD. Indoor allergens and asthma: report of the Third International Workshop. J Allergy Clin Immunol. 1997; 100:S2–S24.4. Neville AC, Parry DA, Woodhead-Galloway J. The chitin crystallite in arthropod cuticle. J Cell Sci. 1976; 21:73–82.5. Fuhrman JA, Piessens WF. Chitin synthesis and sheath morphogenesis in Brugia malayi microfilariae. Mol Biochem Parasitol. 1985; 17:93–104.6. Araujo AC, Souto-Padrón T, de Souza W. Cytochemical localization of carbohydrate residues in microfilariae of Wuchereria bancrofti and Brugia malayi. J Histochem Cytochem. 1993; 41:571–578.7. Debono M, Gordee RS. Antibiotics that inhibit fungal cell wall development. Annu Rev Microbiol. 1994; 48:471–497.8. Jeong KY, Park JW, Hong CS. House dust mite allergy in Korea: the most important inhalant allergen in current and future. Allergy Asthma Immunol Res. 2012; 4:313–325.9. Gao YF, Wang de Y, Ong TC, Tay SL, Yap KH, Chew FT. Identification and characterization of a novel allergen from Blomia tropicalis: Blo t 21. J Allergy Clin Immunol. 2007; 120:105–112.10. Da Silva CA, Pochard P, Lee CG, Elias JA. Chitin particles are multifaceted immune adjuvants. Am J Respir Crit Care Med. 2010; 182:1482–1491.11. Shibata Y, Foster LA, Bradfield JF, Myrvik QN. Oral administration of chitin down-regulates serum IgE levels and lung eosinophilia in the allergic mouse. J Immunol. 2000; 164:1314–1321.12. Strong P, Clark H, Reid K. Intranasal application of chitin microparticles down-regulates symptoms of allergic hypersensitivity to Dermatophagoides pteronyssinus and Aspergillus fumigatus in murine models of allergy. Clin Exp Allergy. 2002; 32:1794–1800.13. Medzhitov R, Janeway C Jr. Innate immunity. N Engl J Med. 2000; 343:338–344.14. Duez C, Gosset P, Tonnel AB. Dendritic cells and toll-like receptors in allergy and asthma. Eur J Dermatol. 2006; 16:12–16.15. Kim YK, Oh SY, Jeon SG, Park HW, Lee SY, Chun EY, et al. Airway exposure levels of lipopolysaccharide determine type 1 versus type 2 experimental asthma. J Immunol. 2007; 178:5375–5382.16. Choi JP, Kim YS, Kim OY, Kim YM, Jeon SG, Roh TY, et al. TNF-alpha is a key mediator in the development of Th2 cell response to inhaled allergens induced by a viral PAMP double-stranded RNA. Allergy. 2012; 67:1138–1148.17. Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. 2010; 11:373–384.18. Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006; 124:783–801.19. Choi JP, Kim YS, Tae YM, Choi EJ, Hong BS, Jeon SG, et al. A viral PAMP double-stranded RNA induces allergen-specific Th17 cell response in the airways which is dependent on VEGF and IL-6. Allergy. 2010; 65:1322–1330.20. Jeon SG, Oh SY, Park HK, Kim YS, Shim EJ, Lee HS, et al. TH2 and TH1 lung inflammation induced by airway allergen sensitization with low and high doses of double-stranded RNA. J Allergy Clin Immunol. 2007; 120:803–812.21. Fuchs B, Braun A. Improved mouse models of allergy and allergic asthma--chances beyond ovalbumin. Curr Drug Targets. 2008; 9:495–502.22. Kumar RK, Herbert C, Foster PS. The "classical" ovalbumin challenge model of asthma in mice. Curr Drug Targets. 2008; 9:485–494.23. Yamashita M, Nakayama T. Progress in allergy signal research on mast cells: regulation of allergic airway inflammation through toll-like receptor 4-mediated modification of mast cell function. J Pharmacol Sci. 2008; 106:332–335.24. Fattouh R, Pouladi MA, Alvarez D, Johnson JR, Walker TD, Goncharova S, et al. House dust mite facilitates ovalbumin-specific allergic sensitization and airway inflammation. Am J Respir Crit Care Med. 2005; 172:314–321.25. Shuhui L, Mok YK, Wong WS. Role of mammalian chitinases in asthma. Int Arch Allergy Immunol. 2009; 149:369–377.26. Reese TA, Liang HE, Tager AM, Luster AD, Van Rooijen N, Voehringer D, et al. Chitin induces accumulation in tissue of innate immune cells associated with allergy. Nature. 2007; 447:92–96.27. Shibata Y, Foster LA, Metzger WJ, Myrvik QN. Alveolar macrophage priming by intravenous administration of chitin particles, polymers of N-acetyl-D-glucosamine, in mice. Infect Immun. 1997; 65:1734–1741.28. Da Silva CA, Chalouni C, Williams A, Hartl D, Lee CG, Elias JA. Chitin is a size-dependent regulator of macrophage TNF and IL-10 production. J Immunol. 2009; 182:3573–3582.29. Da Silva CA, Hartl D, Liu W, Lee CG, Elias JA. TLR-2 and IL-17A in chitin-induced macrophage activation and acute inflammation. J Immunol. 2008; 181:4279–4286.30. Chen Q, Davidson TS, Huter EN, Shevach EM. Engagement of TLR2 does not reverse the suppressor function of mouse regulatory T cells, but promotes their survival. J Immunol. 2009; 183:4458–4466.31. Choi JP, Kim YM, Choi HI, Choi SJ, Park HT, Lee WH, et al. An important role of tumor necrosis factor receptor-2 on natural killer T cells on the development of dsRNA-enhanced Th2 cell response to inhaled allergens. Allergy. 2014; 69:186–198.32. Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol. 2010; 11:889–896.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Innate Immune Response in House Dust Mite-Induced Allergic Inflammation
- House Dust Mite Allergy in Korea: The Most Important Inhalant Allergen in Current and Future
- House dust mite dose not directly activate the human peripheral blood eosinophils in house dust mite-sensitized asthmatics
- Food and house dust mite allergens in children with atopic dermatitis
- Newly sensitization to house dust mite in an isocyanate-induced asthmatic patient
