Evidence for Cyclooxygenase-2 Association with Caveolin-3 in Primary Cultured Rat Chondrocytes
- Affiliations
-
- 1Department of Pharmacology and Toxicology, College of Medicine, Inha University, Incheon, Korea. shcha@inha.ac.kr
- 2Department of Biological Engineering, College of Medicine, Inha University, Incheon, Korea.
- 3National Creative Research Initiative, Center for Secretory Granule Research, College of Medicine, Inha University, Incheon, Korea.
- 4Department of Laboratory Animal, Medical Research Center, College of Medicine, Yonsei University, Seoul, Korea.
- 5Laboratory of Molecular Neuroscience, Department of Life Science, POSTECH, Pohang, Korea.
- KMID: 2157791
- DOI: http://doi.org/10.3346/jkms.2006.21.1.100
Abstract
- The purpose of this study was to demonstrate the cellular localization of cyclooxygenase-2 (COX-2) and caveolin-3 (Cav-3) in primarily cultured rat chondrocytes. In normal rat chondrocytes, we observed relatively high levels of Cav-3 and a very low level of COX-2 mRNA and protein. Upon treating the chondrocytes with 5 microM of CdCl2 (Cd) for 6 hr, the expressions of COX-2 mRNA and protein were increased with the decreased Cav-3 mRNA and protein expressions. The detergent insoluble caveolae-rich membranous fractions that were isolated from the rat chondrocytes and treated with Cd contained the both proteins of both COX-2 and Cav-3 in a same fraction. The immuno-precipitation experiments showed complex formation between the COX-2 and Cav-3 in the rat chondrocytes. Purified COX-2 with glutathione S-transferase-fused COX-2 also showed complex formation with Cav-3. Confocal and electron microscopy also demonstrated the co-localization of COX-2 and Cav-3 in the plasma membrane. The results from our current study show that COX-2 and Cav-3 are co-localized in the caveolae of the plasma membrane, and they form a protein-protein complex. The co-localization of COX-2 with Cav-3 in the caveolae suggests that the caveolins might play an important role for regulating the function of COX-2.
MeSH Terms
-
Animals
Animals, Newborn
Blotting, Western
Cadmium Chloride/pharmacology
Caveolae/drug effects/metabolism/ultrastructure
Caveolin 3/*genetics/metabolism
Cell Membrane/drug effects/metabolism
Cells, Cultured
Chondrocytes/cytology/drug effects/*metabolism
Cyclooxygenase 2/*genetics/metabolism
Gene Expression
Immunoprecipitation
Microscopy, Confocal
Microscopy, Electron
RNA, Messenger/genetics/metabolism
Rats
Reverse Transcriptase Polymerase Chain Reaction
Figure
-

Fig. 1 Morphology and characterization of primarily cultured rat chondrocytes. (A) Light microscopy of confluently grown chondrocytes. (B) Messenger RNA expression of type II collagen and aggrecan in the cultured rat chondrocytes. Total RNA (500 ng) isolated from rat chondrocytes was reverse transcribed and polymerase chain reaction was performed (35 cycles). PCR products were separated using 1% agarose gel and stained with ethidium bromide. (C) Isolated membrane protein (30 µg/lane) was separated by electrophoresis and presence of each type II collagen and aggrecan was detected with respective type II collagen and aggrecan antibodies. Data was shown typical result from 3 independent experiments. M, size marker; C, type II collagen; A, aggrecan.
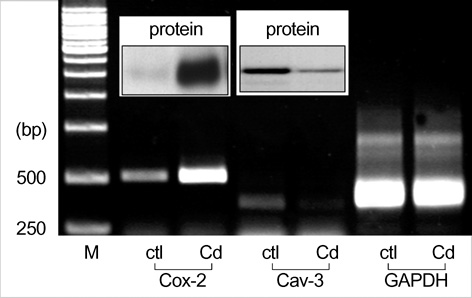
Fig. 2 Expression of COX-2 and Cav-3 mRNA and protein in primarily cultured rat chondrocytes. Cells were changed with medium containing 0.5% FBS in order to exclude serum effect prior to 12 hr of Cd treatment. Then, cells were treated with 5 µM of CdCl2 for 6 hr. After treatment, cells were collected with Tri Reagent for RT-PCR and with lysis buffer for Western blot analysis. The conditions of RT-PCR and Western blotting were same as those of Fig. 1. The COX-2 and Cav-3 antibodies (in Box). Data was shown typical result from 3 independent experiments. ctl; control, Cd; cadmium treated.
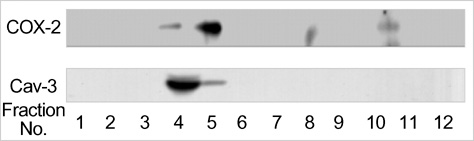
Fig. 3 Western blot analysis of COX-2 and Cav-3 in caveolae-rich membrane fraction. Fractions numbered 4 and 5 are the detergent resistant caveolae-rich fractions obtained by sucrose density gradient centrifugations and the fractions numbered 9 to 12 are the detergent soluble membrane fractions. Fractions obtained from sucrose density gradient centrifugation of detergent solubilized rat chondrocytes were subjected to Western blot analysis with respective antibodies. The result was revealed from 2 separate experiments.

Fig. 4 Complex formation between COX-2 and Cav-3 in the rat chondrocytes. Cell lysates and purified GST fusion proteins (incubated for 2 hr at 37℃ were immunoprecipitated initially with antibodies of COX-2 or Cav-3 and the respective immunoprecipitated (IP) proteins (15 µg) were loaded onto each lane of a 10% SDS-polyacrylamide gel. After separation of the proteins by electrophoresis and transferring the separated proteins to a membrane, the membrane was immunoblotted (IB) using the indicated respective antibodies. Cell lysates were immunoprecipitated initially with COX-2 antibody (A) or Cav-3 antibody (B). The immunoprecipitational result for GST fusion proteins was shown in (C).
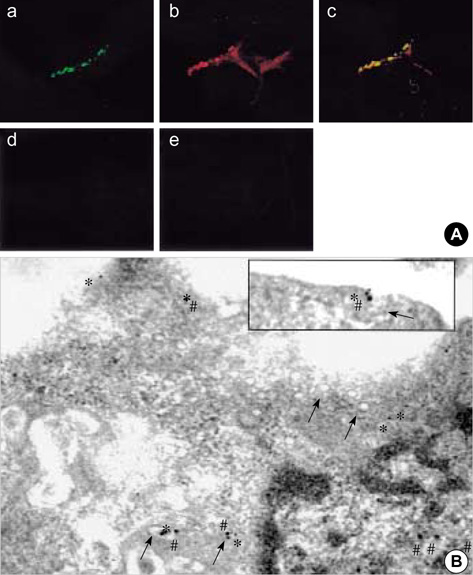
Fig. 5 Confocal and electron microscopy of rat chondrocytes immuno-cytochemically stained with COX-2 and caveolin antibodies. Cells were double stained with fluorescein isothiocyanate- and Texas Red-conjugated secondary antibodies after an initial treatment with either Cav-3 or COX-2 antibodies separately and then examined under a confocal microscope at their respective wavelengths. For each experiment, a least 50 cells were examined and the presented images represent typical staining pattern for the majority of examined cells. a, Cav-3; b, COX-2; c, merged image of a and b; d and e; stained only with secondary antibodies without the prior treatment of either Cav-3 or COX-2 antibodies (A). Cells were double stained with different size of gold particle-conjugated secondary antibodies after an initial treatment with either Cav-3 or COX-2 antibodies separately and then examined under an electron microscope. Arrow showed structure of caveolae. # and * represented COX-2 (15 nm particles) and Cav-3 (10 nm particles), respectively.
Reference
-
1. Nasrallah R, Hebert RL. Prostacyclin signaling in the kidney: implications for health and disease. Am J Physiol. 2005. 289:235–246.
Article2. Smith WL, Garavito RM, DeWitt DL. Prostaglandin endoperoxide H synthases (cyclooxygenase)-1 and -2. J Biol Chem. 1996. 271:33157–33160.3. Rodrigues S, Bruyneel E, Rodrigue CM, Shahin E, Gespach C. Cyclooxygenase 2 and carcinogenesis. Bull Cancer. 2004. 91:S61–S76.4. Park MK, Hwang SY, Kim JO, Kwack MH, Kim JC, Kim MK, Sung YK. NS398 inhibits the growth of Hep3B human hepatocellular carcinoma cells via caspase-independent apoptosis. Mol Cells. 2004. 17:45–50.5. O'Neill GP, Ford-Hutchinson AW. Expression of mRNA for cyclooxygenase-2 and cyclooxygenase-2 in human tissues. FEBS Lett. 1993. 330:156–160.6. Wu KK. Cyclooxygenase 2 induction: molecular mechanism and pathophysiologic roles. J Lab Clin Med. 1996. 128:242–245.
Article7. Williams TM, Lisanti MP. The caveolin genes: from cell biology to medicine. Ann Med. 2004. 36:584–595.
Article8. Liu P, Rudick M, Anderson RG. Multiple functions of caveolin-1. J Biol Chem. 2002. 277:41295–41298.
Article9. Liou JY, Deng WG, Gilroy DW, Shyue SK, Wu KK. Colocalization and interaction of cyclooxygenase-2 with caveolin-1 in human fibroblasts. J Biol Chem. 2001. 276:34975–34982.
Article10. Cha SH, Jung NH, Kim BR, Kim HW, Kwak JO. Evidence for cyclooxygenase-1 association with caveolin-1 and -2 in cultured human embryonic kidney (HEK 293) cells. IUBMB Life. 2004. 56:221–227.11. Ikezu T, Ueda H, Trapp BD, Nishiyama K, Sha JF, Volonte D, Galbiati F, Byrd AL, Bassell G, Serizawa H, Lane WS, Lisanti MP, Okamoto T. Affinity-purification and characterization of caveolins from the brain: differential expression of caveolin-1, -2, and -3 in brain endothelial and astroglial cell types. Brain Res. 1998. 804:177–192.
Article12. Schwab W, Galbiati F, Volonte D, Hempel U, Wenzel KW, Funk RH, Lisanti MP, Kasper M. Characterisation of caveolins from cartilage: expression of caveolin-1, -2 and -3 in chondrocytes and in alginate cell culture of the rat tibia. Histochem Cell Biol. 1999. 112:41–49.
Article13. Kiepe D, Andress DL, Mohan S, Standker L, Ulinski T, Himmele R, Mehls O, Tonshoff B. Intact IGF-binding protein-4 and -5 and their respective fragments isolated from chronic renal failure serum differentially modulate IGF-1 actions in cultured growth plate chondrocytes. J Am Soc Nephrol. 2001. 12:2400–2410.14. Ferretti A, Knijn A, Raggi C, Sargiacomo M. High-resolution proton NMR measures mobile lipids associated with triton-resistant membrane domains in haematopoietic K562 cells lacking or expression caveolin-1. Eur Biophys J. 2003. 32:83–95.15. Spector DL, Fu XD, Maniatis T. Associations between distinct premRNA splicing components and the cell nucleus. EMBO J. 1991. 10:3467–3481.
Article16. Harris ED Jr. Rheumatoid arthritis. Pathophysiology and implications for therapy. N Engl J Med. 1990. 322:1277–1289.17. Hardy MM, Seibert K, Manning PT, Currie MG, Woerner BM, Edwards D, Koki A, Tripp CS. Cyclooxygenase 2-dependent prostaglandin E2 modulates cartilage proteoglycan degradation in human osteoarthritis explants. Arthritis Rheum. 2002. 46:1789–1803.
Article18. Blanco FJ, Guitian R, Moreno J, de Toro FJ, Galdo F. Effect of anti-inflammatory drugs on COX-1 and COX-2 activity in human articular chondrocytes. J Rheumatol. 1999. 26:1366–1373.19. Choi YA, Lee DJ, Lim HK, Jeong JH, Sonn JK, Kang SS, Baek SH. Interleukin-1β stimulates matrix metalloproteinase-2 expression via a prostaglandin E2-dependent mechanism in human chondrocytes. Exp Mol Med. 2004. 36:226–232.20. Gilroy DW, Colville-Nash PR, Willis D, Chivers J, Paul-Clark MJ, Willoughby DA. Inducible cyclooxygenase may have anti-inflammatory properties. Nat Med. 1999. 5:698–701.
Article21. Fahmi H, Pelletier JP, Martel-Pelletier J. PPARgamma ligands as modulators of inflammatory and catabolic responses in arthritis. An overview. J Rheumatol. 2002. 29:3–14.22. Sandell LJ, Adler P. Developmental patterns of cartilage. Front Biosci. 1999. 4:731–742.
Article23. DeLise AM, Fischer L, Tuan RS. Cellular interactions and signaling in cartilage development. Osteoarthritis Cartilage. 2000. 8:309–334.
Article24. Li XA, Everson WV, Smart EJ. Caveolae, lipid rafts, and vascular disease. Trends Cardiovasc Med. 2005. 15:92–96.
Article25. Couet J, Li S, Okamoto T, Ikezu T, Lisanti MP. Identification of peptide and protein ligands for the caveolin-scaffolding domain. Implications for the interaction of caveolin with caveolae-associated proteins. J Biol Chem. 1997. 272:6525–6533.26. Parton RG. Caveolae and caveolins. Curr Opin Cell Biol. 1996. 8:542–548.
Article27. Okamoto T, Schlegel A, Scherer PE, Lisanti MP. Caveolins, a family of scaffolding proteins for organizing "preassembled signaling complexes" at the plasma membrane. J Biol Chem. 1998. 273:5419–5422.
Article28. Scherer PE, Lewis RY, Volonte D, Engelman JA, Galbiati F, Couet J, Kohtz DS, Donselaar E, Perters P, Lisanti MP. Cell-type and tissue-specific expression of caveolin-2: Caveolins 1 and 2 co-localize and form a stable hetero-oligomeric complex in vivo. J Biol Chem. 1997. 272:29337–29346.29. Li S, Galbiati F, Volonte D, Sargiacomo M, Engelman JA, Das K, Scherer PE, Lisanti MP. Mutational analysis of caveolin-induced vesicle formation. Expression of caveolin-1 recruits caveolin-2 to caveolae membranes. FEBS Lett. 1998. 434:127–134.30. Song KS, Scherer PE, Tang Z, Okamoto T, Li S, Chafel M, Chu C, Kohtz DS, Lisanti MP. Expression of Caveolin-3 in Skeletal, Cardiac, and Smooth Muscle Cells. Caveolin-3 is a component of the sarcolemma and co-fractionates with dystrophin and dystrophin-associated glycoproteins. J Biol Chem. 1996. 271:15160–15165.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Ectopic Expression of Caveolin-1 Induces COX-2 Expression in Rabbit Articular Chondrocytes via MAP Kinase Pathway
- Src Kinase Regulates Nitric Oxide-induced Dedifferentiation and Cyclooxygenase-2 Expression in Articular Chondrocytes via p38 Kinase-dependent Pathway
- p38 Kinase Regulates Nitric Oxide-induced Dedifferentiation and Cyclooxygenase-2 Expression of Articular Chondrocytes
- Expression of Caveolin-3 in the Myelin Sheath of Peripheral Nerve
- Immunohistochemical study of caveolin-1 and -2 in the rat retina
