MLL4 Regulates the Progression of Non–Small-Cell Lung Cancer by Regulating the PI3K/AKT/SOX2 Axis
- Yang Y
 1,2,3
1,2,3 - Qiu R1,2,3
- Weng Q1,2,3
- Xu Z1,2,3
- Song J1,2,3
- Zhao S1,2,3
- Meng M1,2,3
- Zhang D1,2,3
- Kong C1,2,3
- Wang H1,2,3
- Xu M1,2,3
- Zhao Z1,2,3
- Ji J1,2,3
- Affiliations
-
- 1Key Laboratory of Imaging Diagnosis and Minimally Invasive Intervention Research, Institute of Imaging Diagnosis and Minimally Invasive Intervention Research, The Fifth Affiliated Hospital of Wenzhou Medical University, Lishui, China
- 2Department of Radiology, Clinical College of The Affiliated Central Hospital, Lishui University, Lishui, China
- 3Department of Radiology, Lishui Hospital of Zhejiang University, Lishui, China
- KMID: 2544161
- DOI: http://doi.org/10.4143/crt.2022.1042
Abstract
- Purpose
Mixed-lineage leukemia protein 4 (MLL4/KMT2D) is a histone methyltransferase, and its mutation has been reported to be associated with a poor prognosis in many cancers, including lung cancer. We investigated the function of MLL4 in lung carcinogenesis.
Materials and Methods
RNA sequencing (RNA-seq) in A549 cells transfected with control siRNA or MLL4 siRNA was performed. Also, we used EdU incorporation assay, colony formation assays, growth curve analysis, transwell invasion assays, immunohistochemical staining, and in vivo bioluminescence assay to investigate the function of MLL4 in lung carcinogenesis.
Results
We found that MLL4 expression was downregulated in non–small cell lung cancer (NSCLC) tissues compared to adjacent normal tissues and tended to decrease with disease stage progression. We analyzed the transcriptomes in control and MLL4- deficient cells using high-throughput RNA deep sequencing (RNA-seq) and identified a cohort of target genes, such as SOX2, ATF1, FOXP4, PIK3IP1, SIRT4, TENT5B, and LFNG, some of which are related to proliferation and metastasis. Our results showed that low expression of MLL4 promotes NSCLC cell proliferation and metastasis and is required for the maintenance of NSCLC stem cell properties.
Conclusion
Our findings identify an important role of MLL4 in lung carcinogenesis through transcriptional regulation of PIK3IP1, affecting the PI3K/AKT/SOX2 axis, and suggest that MLL4 could be a potential prognostic indicator and target for NSCLC therapy.
Keyword
Figure
-

Fig. 1 Mixed-lineage leukemia protein 4 (MLL4) expression is downregulated in non–small cell lung cancer (NSCLC) tissue. (A) Analysis of public datasets (GSE33356, GSE19188, and GSE18442) for the expression of MLL4 by two-tailed unpaired t test. (B) MLL4 alteration frequency from five lung cancer data sets, data analysis was performed on the ciBioportal platform. LUAD, lung adenocarcinoma; LUSC, lung squamous cell carcinoma; MSKCC, Memorial Sloan Kettering Cancer Center; SCLC, small cell lung cancer; TCGA, The Cancer Genome Atlas. (C) Real-time polymerase chain reaction (PCR) of MLL4 in 16 pairs of NSCLC and adjacent normal tissues. The expression of MLL4 was measured by quantitative real-time PCR. In addition, mRNA levels were normalized to that of β-actin. Each bar represents the mean±standard deviation for triplicate experiments. (D) Immunohistochemical staining of MLL4 in 16 pairs of NSCLC and adjacent normal tissues. For each group, representative photos of two specimens are shown. (E) H&E staining and immunohistochemical staining of MLL4 in 35 pairs of NSCLC and adjacent normal tissues with TNM clinical classification (adjacent, stage 0, I, II, III, and IV). For each stage, representative photos of two specimens are shown. (F, G) The positively stained nuclei (%) in 35 paired samples were analyzed by two-tailed unpaired t test (*p < 0.05, **p < 0.01, ***p < 0.001).

Fig. 2 Genome-wide identification of transcription targets for mixed-lineage leukemia protein 4 (MLL4). (A) A549 cells were transfected with control siRNA and MLL4 siRNA followed by RNA extraction and deep sequencing. (B) Gene set enrichment analysis (GSEA) results showing enriched gene signatures related to the transforming growth factor β (TGF-β) signaling pathway in A549 cells infected with control siRNA or MLL4 siRNA. (C) MLL4-regulated genes were grouped and statistically analyzed according to Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways. (D) The heatmap presented the effect of MLL4 knockdown on the expression of genes. (E, F) A549 cells or SK-MES-1 cells were transfected with the indicated siRNAs, followed by RNA extraction and quantitative reverse transcription PCR analysis of the expression of the indicated genes. The mRNA levels were normalized to those of β-actin. *p < 0.05, **p < 0.01, and ***p < 0.001 (two-tailed t test). (G) A549 cells or SK-MES-1 cells were transfected with the indicated siRNAs, followed by western blot analysis of the expression of the indicated genes. (H) A549 cells or SK-MES-1 cells were transfected with MLL4 siRNA #2, and the protein levels of H3K4me1 were measured. H3 served as loading control for the western blot. (I) A549 cells transfected with indicated siRNAs and plasmid and Western blot analysis of indicated target genes. The knockdown efficiencies of MLL4 were confirmed by quantitative real-time PCR. (J) Control siRNA or MLL4 siRNA transfected A549 and SK-MES-1 cells were treated in the presence or absence of PI3K inhibitor (ZSTK474) at 10 μm concentration for 24 hours. The p-AKT and SOX2 levels are shown. β-Actin was used as a loading control. AGE-RAGE, advanced glycation end-products–receptor for advanced glycation end products; ECM, extracellular matrix; IL-17, interleukin 17; PI3K, phosphoinositide 3-kinase; TGF, transforming growth factor; TNF, tumor necrosis factor.

Fig. 3 Stable inhibition of mixed-lineage leukemia protein 4 (MLL4) promotes the proliferation in non–small cell lung cancer cells. (A, B) A549 cells and SK-MES-1 cells were transfected with CRISPR activation (CRISPRa)-Scramble or CRISPRa-MLL4 gRNA or the indicated specific shRNAs or/and expression constructs for EdU assays. (C, D) A549 cells and SK-MES-1 cells stably infected with CRISPRa-Scramble or CRISPRa-MLL4 gRNA or the indicated specific shRNAs or/and expression constructs in culture media for 10 days prior to being stained with crystal violet. Representative photos and statistical analyses are shown. (E) The activation and knockdown efficiencies of MLL4 were confirmed by quantitative real-time PCR. The overexpression of PIK3IP1 was confirmed by western blot analysis. Each bar represents the mean±standard deviation for triplicate measurements. *p < 0.05 and **p < 0.01 (two-tailed t test).

Fig. 4 The effect of PIK3IP1 and SOX2 on non–small cell lung cancer cells proliferation. (A, B) A549 cells and SK-MES-1 cells were transfected with the indicated specific shRNAs or/and expression constructs for EdU assays. (C, D) A549 cells and SK-MES-1 cells stably infected with the indicated specific shRNAs or/and expression constructs in culture media for 10 days prior to being stained with crystal violet. (E) Western blot analysis was used to determine the protein expression in these cells using antibodies against the indicated proteins. Each bar represents the mean±standard deviation for triplicate measurements. **p < 0.01 (two-tailed t test).

Fig. 5 Knockdown of mixed-lineage leukemia protein 4 (MLL4) promotes the invasion and migration of non–small cell lung cancer cells. (A, B) A549 and SK-MES-1 cells were transfected with the indicated specific shRNAs or/and expression constructs subjected to growth curve analysis by counting the numbers of living cells. (C, D) A549 cells and SK-MES-1 cells were transfected with Scramble or MLL4 gRNA or the indicated specific shRNAs or/and expression constructs. The migration of cells towards the wound was photographed under a light microscope. Representative photos and statistical analyses are shown. (E, F) Transwell invasion assays of A549 and SK-MES-1 cells following transfection with CRISPR activation (CRISPRa)-Scramble or CRISPRa-MLL4 gRNA or the indicated specific shRNAs and/or expression constructs. **p < 0.01 and ***p < 0.001 (two-tailed t test). (G) The activation and knockdown efficiencies of MLL4 were confirmed by quantitative real-time polymerase chain reaction. The overexpression of PIK3IP1 was confirmed by western blot analysis.

Fig. 6 The effect of PIK3IP1 and SOX2 on non–small cell lung cancer cells migration and invasion. (A, B) A549 cells and SK-MES-1 cells were transfected with the indicated specific shRNAs or/and expression constructs. The migration of cells towards the wound was photographed under a light microscope. (C, D) Transwell invasion assays of A549 and SK-MES-1 cells following transfection with the indicated specific shRNAs or/and expression constructs. The invaded cells were stained and counted. (E) Western blot analysis was used to determine the protein expression in these cells using antibodies against the indicated proteins. Each bar represents the mean±standard deviation for triplicate measurements. *p < 0.05, **p < 0.01, and ***p < 0.001 (two-tailed t test).

Fig. 7 Stable inhibition of mixed-lineage leukemia protein 4 (MLL4) promotes sphere formation of non–small cell lung cancer cells. A549 (A) and SK-MES-1 cells (B) transfected with CRISPR activation (CRISPRa)-Scramble or CRISPRa-MLL4 gRNA or the indicated specific shRNAs or/and expression constructs were plated in the ultra-low attachment 24-well plate at a density of 1,000 cells/well and cultured in tumor sphere medium. Tumor spheres were photographed and counted. **p < 0.01 (two-tailed t test). (C) The activation and knockdown efficiencies of MLL4 were confirmed by quantitative real-time polymerase chain reaction. The overexpression of PIK3IP1 were confirmed by western blot analysis.
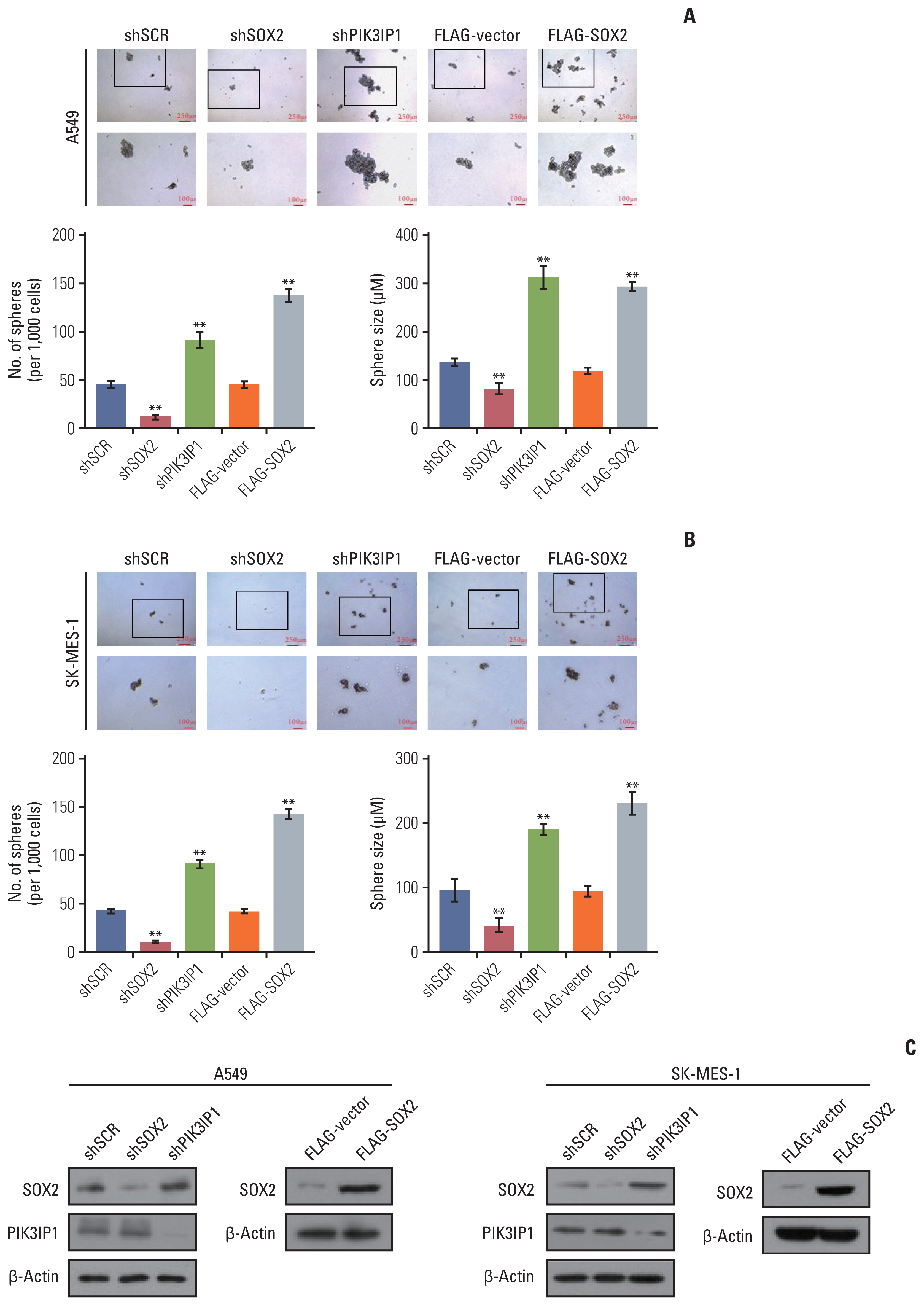
Fig. 8 The effect of PIK3IP1 and SOX2 on non–small cell lung cancer cells sphere formation ability. (A, B) A549 and SK-MES-1 cells transfected with the indicated specific shRNAs or/and expression constructs were performed Sphere Formation Assays. Tumor spheres were photographed and counted. (C) Western blot analysis was used to determine the protein expression in these cells using antibodies against the indicated proteins. Each bar represents the mean±standard deviation for triplicate measurements. **p < 0.01 (two-tailed t test).

Fig. 9 Stable inhibition of mixed-lineage leukemia protein 4 (MLL4) promotes tumorigenesis in vivo. (A) A549 cells stably expressing MLL4 shRNA or shSCR were transplanted into athymic mice. Representative images of tumor-bearing mice and their tumors, the average tumor mass, and weight of each group are shown. **p < 0.01 (two-tailed unpaired t test). (B) A549 cells that had been engineered to stably express firefly luciferase (A549-Luc) were infected with lentiviruses carrying shRNA against MLL4 and then injected intravenously through the tail vein of 6-week-old male NOD SCID mice (n=5). Lung metastasis was quantified using bioluminescence imaging after 4 weeks. Representative in vivo bioluminescent images are shown. Lung cancer specimens were examined by in vitro bioluminescent measurement and the sections from control or shMLL4-treated mice were stained with H&E. (C) Kaplan-Meier survival analysis for the relationship between survival time and the MLL4, PIK3IP1, and SOX2 signature in lung cancer using an online tool (http://kmplot.com/analysis). HR, hazard ratio. (D) Analysis of the correlations between MLL4 and PIK3IP1 and SOX2 in the public datasets GSE108492, GSE115457, GSE8894, and GSE3141.
Reference
-
References
1. Maiuthed A, Chantarawong W, Chanvorachote P. Lung cancer stem cells and cancer stem cell-targeting natural compounds. Anticancer Res. 2018; 38:3797–809.
Article2. Yang S, Zhang Z, Wang Q. Emerging therapies for small cell lung cancer. J Hematol Oncol. 2019; 12:47.
Article3. Dong S, Li W, Wang L, Hu J, Song Y, Zhang B, et al. Histone-related genes are hypermethylated in lung cancer and hypermethylated HIST1H4F could serve as a pan-cancer biomarker. Cancer Res. 2019; 79:6101–12.4. Fagan RJ, Dingwall AK. COMPASS ascending: emerging clues regarding the roles of MLL3/KMT2C and MLL2/KMT-2D proteins in cancer. Cancer Lett. 2019; 458:56–65.
Article5. Gonda TJ, Ramsay RG. Directly targeting transcriptional dysregulation in cancer. Nat Rev Cancer. 2015; 15:686–94.6. Ford DJ, Dingwall AK. The cancer COMPASS: navigating the functions of MLL complexes in cancer. Cancer Genet. 2015; 208:178–91.7. Ardeshir-Larijani F, Bhateja P, Lipka MB, Sharma N, Fu P, Dowlati A. KMT2D mutation is associated with poor prognosis in non-small-cell lung cancer. Clin Lung Cancer. 2018; 19:e489–501.
Article8. Lin-Shiao E, Lan Y, Coradin M, Anderson A, Donahue G, Simpson CL, et al. KMT2D regulates p63 target enhancers to coordinate epithelial homeostasis. Genes Dev. 2018; 32:181–93.9. Froimchuk E, Jang Y, Ge K. Histone H3 lysine 4 methyltransferase KMT2D. Gene. 2017; 627:337–42.10. Ruthenburg AJ, Allis CD, Wysocka J. Methylation of lysine 4 on histone H3: intricacy of writing and reading a single epigenetic mark. Mol Cell. 2007; 25:15–30.11. Dawkins JB, Wang J, Maniati E, Heward JA, Koniali L, Kocher HM, et al. Reduced expression of histone methyltransferases KMT2C and KMT2D correlates with improved outcome in pancreatic ductal adenocarcinoma. Cancer Res. 2016; 76:4861–71.12. Lv S, Ji L, Chen B, Liu S, Lei C, Liu X, et al. Histone methyltransferase KMT2D sustains prostate carcinogenesis and metastasis via epigenetically activating LIFR and KLF4. Oncogene. 2018; 37:1354–68.
Article13. Hillman RT, Celestino J, Terranova C, Beird HC, Gumbs C, Little L, et al. KMT2D/MLL2 inactivation is associated with recurrence in adult-type granulosa cell tumors of the ovary. Nat Commun. 2018; 9:2496.
Article14. Sun P, Wu T, Sun X, Cui Z, Zhang H, Xia Q, et al. KMT2D inhibits the growth and metastasis of bladder cancer cells by maintaining the tumor suppressor genes. Biomed Pharmacother. 2019; 115:108924.15. Ortega-Molina A, Boss IW, Canela A, Pan H, Jiang Y, Zhao C, et al. The histone lysine methyltransferase KMT2D sustains a gene expression program that represses B cell lymphoma development. Nat Med. 2015; 21:1199–208.
Article16. Xiong W, Deng Z, Tang Y, Deng Z, Li M. Downregulation of KMT2D suppresses proliferation and induces apoptosis of gastric cancer. Biochem Biophys Res Commun. 2018; 504:129–36.
Article17. Alam H, Tang M, Maitituoheti M, Dhar SS, Kumar M, Han CY, et al. KMT2D deficiency impairs super-enhancers to confer a glycolytic vulnerability in lung cancer. Cancer Cell. 2020; 37:599–617.
Article18. Zeng Y, Qiu R, Yang Y, Gao T, Zheng Y, Huang W, et al. Regulation of EZH2 by SMYD2-mediated lysine methylation is implicated in tumorigenesis. Cell Rep. 2019; 29:1482–98.
Article19. Yang Y, Qiu R, Zhao S, Shen L, Tang B, Weng Q, et al. SMYD3 associates with the NuRD (MTA1/2) complex to regulate transcription and promote proliferation and invasiveness in hepatocellular carcinoma cells. BMC Biol. 2022; 20:294.
Article20. Sikorska M, Sandhu JK, Deb-Rinker P, Jezierski A, Leblanc J, Charlebois C, et al. Epigenetic modifications of SOX2 enhancers, SRR1 and SRR2, correlate with in vitro neural differentiation. J Neurosci Res. 2008; 86:1680–93.21. Ediriweera MK, Tennekoon KH, Samarakoon SR. Role of the PI3K/AKT/mTOR signaling pathway in ovarian cancer: biological and therapeutic significance. Semin Cancer Biol. 2019; 59:147–60.
Article22. Tewari D, Patni P, Bishayee A, Sah AN, Bishayee A. Natural products targeting the PI3K-Akt-mTOR signaling pathway in cancer: a novel therapeutic strategy. Semin Cancer Biol. 2022; 80:1–17.
Article23. Yang Q, Jiang W, Hou P. Emerging role of PI3K/AKT in tumor-related epigenetic regulation. Semin Cancer Biol. 2019; 59:112–24.
Article24. Wang Z, Kang L, Zhang H, Huang Y, Fang L, Li M, et al. AKT drives SOX2 overexpression and cancer cell stemness in esophageal cancer by protecting SOX2 from UBR5-mediated degradation. Oncogene. 2019; 38:5250–64.
Article25. Mamun MA, Mannoor K, Cao J, Qadri F, Song X. SOX2 in cancer stemness: tumor malignancy and therapeutic potentials. J Mol Cell Biol. 2020; 12:85–98.
Article26. Karachaliou N, Rosell R, Viteri S. The role of SOX2 in small cell lung cancer, lung adenocarcinoma and squamous cell carcinoma of the lung. Transl Lung Cancer Res. 2013; 2:172–9.27. Singh S, Trevino J, Bora-Singhal N, Coppola D, Haura E, Altiok S, et al. EGFR/Src/Akt signaling modulates Sox2 expression and self-renewal of stem-like side-population cells in non-small cell lung cancer. Mol Cancer. 2012; 11:73.
Article28. Schaefer T, Lengerke C. SOX2 protein biochemistry in stem-ness, reprogramming, and cancer: the PI3K/AKT/SOX2 axis and beyond. Oncogene. 2020; 39:278–92.29. Schaefer T, Ramadoss A, Leu S, Tintignac L, Tostado C, Bink A, et al. Regulation of glioma cell invasion by 3q26 gene products PIK3CA, SOX2 and OPA1. Brain Pathol. 2019; 29:336–50.
Article30. Tang J, Zhong G, Wu J, Chen H, Jia Y. SOX2 recruits KLF4 to regulate nasopharyngeal carcinoma proliferation via PI3K/AKT signaling. Oncogenesis. 2018; 7:61.
Article31. Ormsbee Golden BD, Wuebben EL, Rizzino A. Sox2 expression is regulated by a negative feedback loop in embryonic stem cells that involves AKT signaling and FoxO1. PLoS One. 2013; 8:e76345.
Article32. Egolf S, Zou J, Anderson A, Simpson CL, Aubert Y, Prouty S, et al. MLL4 mediates differentiation and tumor suppression through ferroptosis. Sci Adv. 2021; 7:eabj9141.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Gallic Acid Hindered Lung Cancer Progression by Inducing Cell Cycle Arrest and Apoptosis in A549 Lung Cancer Cells via PI3K/Akt Pathway
- Dual Inhibition of PI3K/Akt/mTOR Pathway and Role of Autophagy in Non-Small Cell Lung Cancer Cells
- Membrane Trafficking of Collecting Duct Water Channel Protein AQP2 Regulated by Akt/AS160
- Up-regulation of P13K/Akt Signaling by 17 beta-estradiol through Activation of Estrogen Receptor-alpha in Breast Cancer Cells
- Long Noncoding RNA PVT1 Promotes Stemness and Temozolomide Resistance through miR-365/ELF4/SOX2 Axis in Glioma
