BRAFV600E Mutation Enhances Estrogen-Induced Metastatic Potential of Thyroid Cancer by Regulating the Expression of Estrogen Receptors
- Affiliations
-
- 1Cancer Research Institute, Seoul National University College of Medicine, Seoul, Korea
- 2Department of Human Genetics, McGill University, Montreal, QC, Canada
- 3Department of Surgery, Seoul National University College of Medicine, Seoul, Korea
- 4Division of Surgery, Thyroid Center, Seoul National University Hospital, Seoul, Korea
- 5Department of Surgery, Yanbian University Hospital, Yanji, China
- 6Department of Internal Medicine, Seoul National University College of Medicine, Seoul, Korea
- KMID: 2537289
- DOI: http://doi.org/10.3803/EnM.2022.1563
Abstract
- Background
Cross-talk between mitogen-activated protein kinase and estrogen has been reported; however, the role of BRAFV600E in the estrogen responsiveness of thyroid cancer is unknown. We elucidated the effect of BRAFV600E on the estrogen-induced increase in metastatic potential in thyroid cancer.
Methods
Using a pair of cell lines, human thyroid cell lines which harbor wild type BRAF gene (Nthy/WT) and Nthy/BRAFV600E (Nthy/V600E), the expression of estrogen receptors (ERs) and estrogen-induced metastatic phenotypes were evaluated. Susceptibility to ERα- and ERβ-selective agents was evaluated to confirm differential ER expression. ESR expression was analyzed according to BRAFV600E status and age (≤50 years vs. >50 years) using The Cancer Genome Atlas (TCGA) data.
Results
Estradiol increased the ERα/ERβ expression ratio in Nthy/V600E, whereas the decreased ERα/ERβ expression ratio was found in Nthy/WT. BRAFV600E-mutated cell lines showed a higher E2-induced increase in metastatic potential, including migration, invasion, and anchorage-independent growth compared with Nthy/WT. An ERα antagonist significantly inhibited migration in Nthy/V600E cells, whereas an ERβ agonist was more effective in Nthy/WT. In the BRAFV600E group, ESR1/ESR2 ratio was significantly higher in younger age group (≤50 years) compared with older age group (>50 years) by TCGA data analysis.
Conclusion
Our data show that BRAFV600E mutation plays a crucial role in the estrogen responsiveness of thyroid cancer by regulating ER expression. Therefore, BRAFV600E might be used as a biomarker when deciding future hormone therapies based on estrogen signaling in thyroid cancer patients.
Figure
-
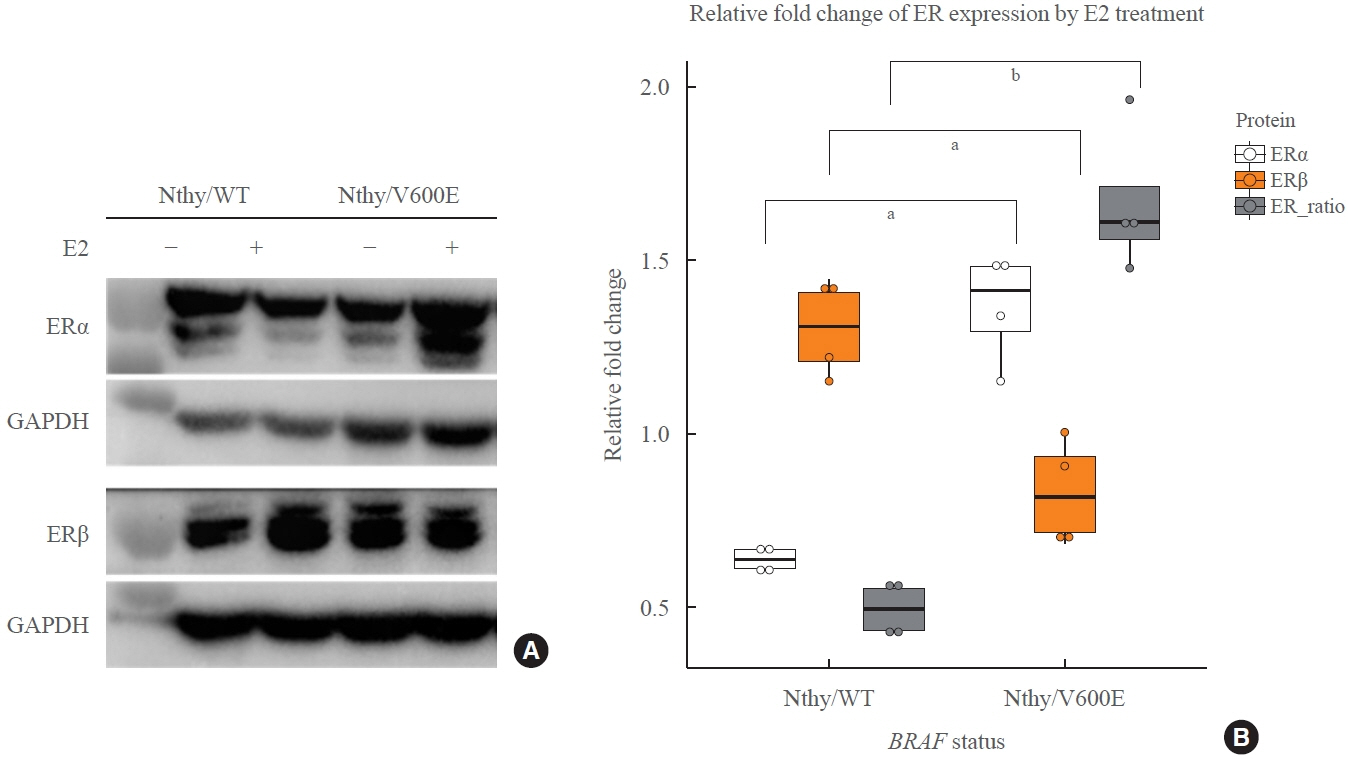
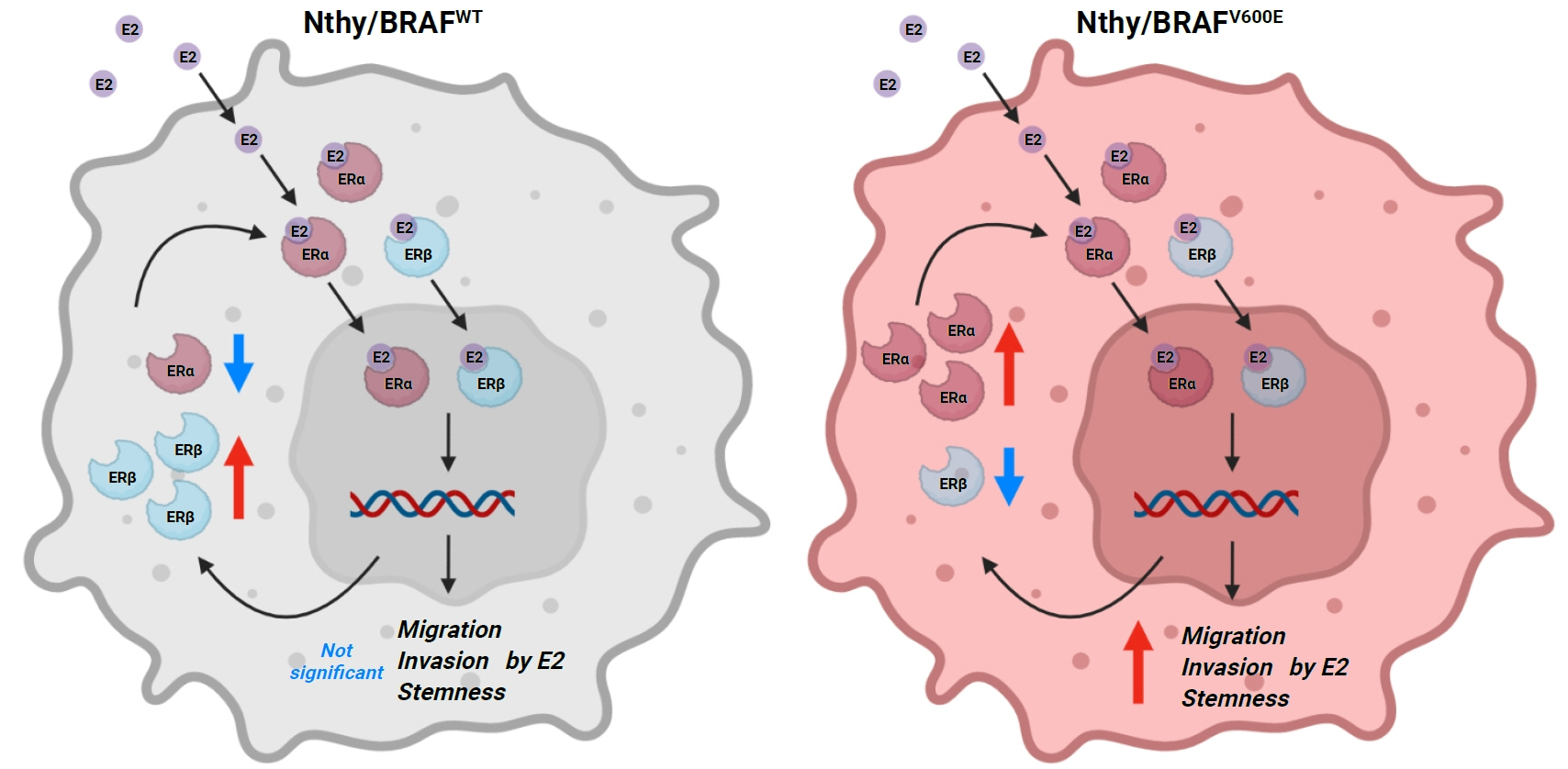
Fig. 1. Expression change of estrogen receptor α (ERα) and ERβ in thyroid cell lines by E2 treatment depending on BRAFV600E status. (A) Expression of estrogen receptors were measured to see how BRAFV600E affects regulation of the receptors in thyroid cell lines. All cell lines were cultured in phenol red-free complete medium and treated with 100 nM of E2 for 24 hours. The response to E2 treatment was opposite between human thyroid cell lines which harbor wild type BRAF gene (Nthy/WT) and Nthy/BRAFV600E (Nthy/V600E). It shows increased ERα and decreased ERβ in Nthy/V600E whereas decreased ERα and increased ERβ in Nthy/WT by E2 treatment. (B) Densitometry results for fold change of ERα, ERβ, and the ratio of ERα over ERβ after E2 treatment. Each value was calculated by fold change of glyceraldehyde-3-phosphate dehydrogenase (GAPDH)-normalized levels of ER proteins on the same membranes. E2 treatment induced decreased ER ratio in Nthy/WT, but increased in Nthy/V600E (0.50 vs. 1.67, P=0.0006). aP<0.01; bP<0.001.
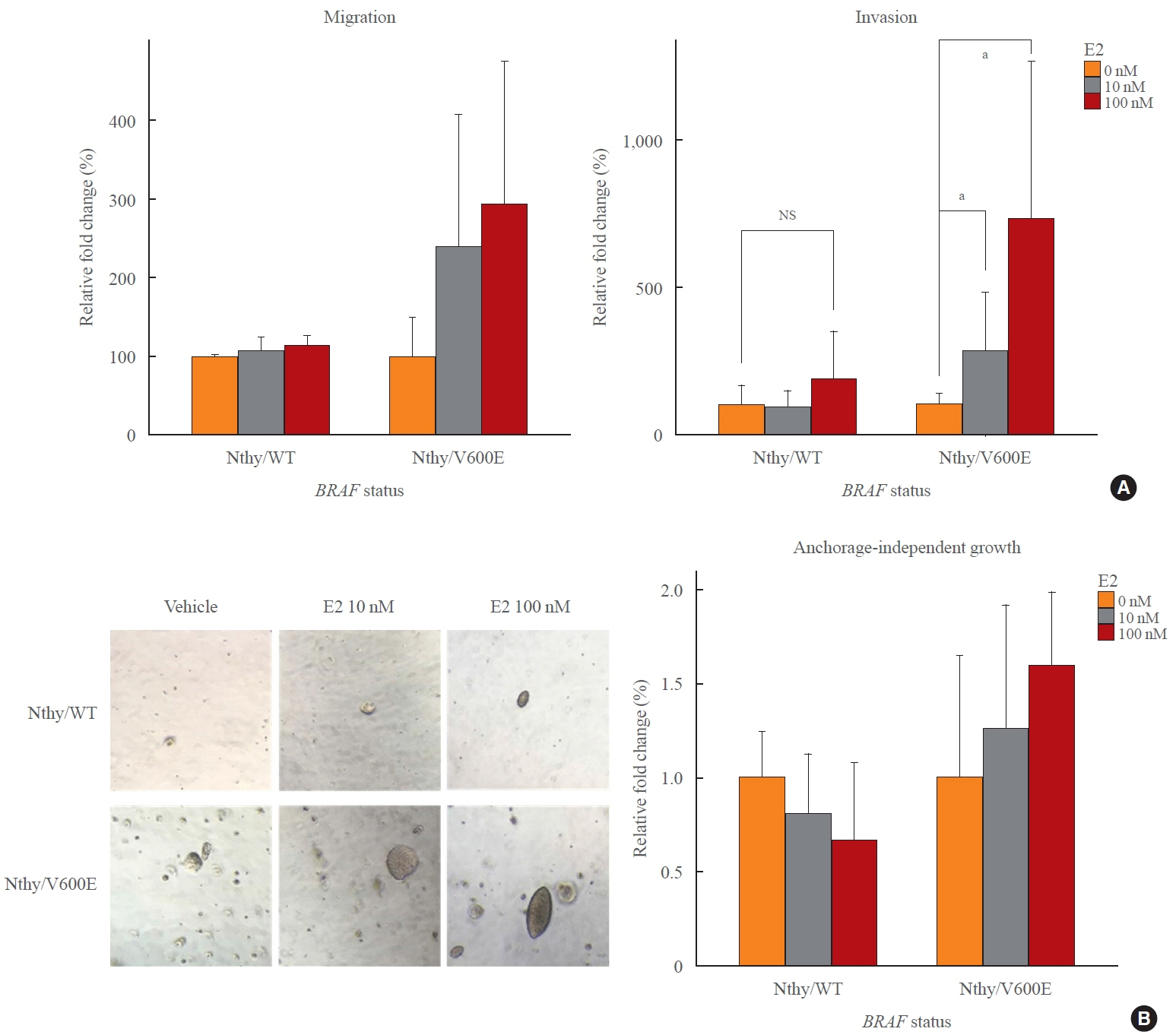
Fig. 2. Enhanced increase of metastatic potentials of thyroid cell lines by BRAFV600E. (A) Human thyroid cell lines which harbor wild type BRAF gene (Nthy/WT) did not show significant changes of migration and invasion by E2. However, BRAFV600E-positive cell lines, and Nthy/BRAFV600E (Nthy/V600E), showed over two times more increase of migration and significant increase of invasion in response to E2 treatment. (B) Anchorage-independent growth of thyroid cell lines was observed under microscopy and the total amount of cells was measured using fluorescent dye included in the soft-agar assay kit. While E2 slightly inhibited growth of Nthy/WT, it promoted colony formation and growth in BRAFV600E-positive cell lines in a dose-dependent manner. NS, not significant. aP<0.05.
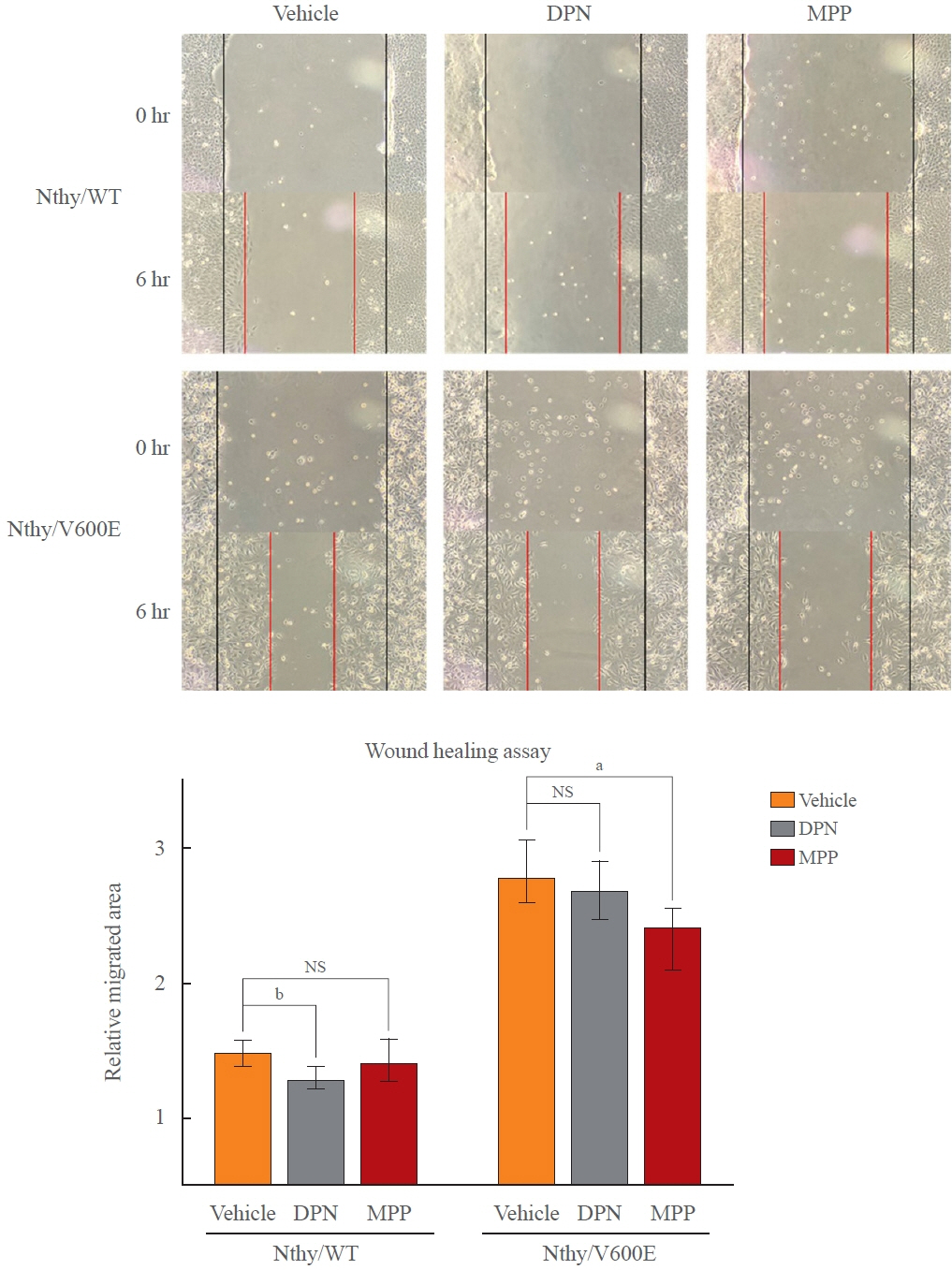
Fig. 3. Altered anti-migratory effects of selective estrogen receptor α (ERα) antagonist and ERβ agonist. Wound healing assay was used to see if susceptibilities to diarylpropionitrile (DPN) and methylpiperidino pyrazole (MPP), which activate ERβ and block ERα, respectively. Both result in inhibition of cell migration. Under presence of endogenous estrogen and estrogenic phenol red, treatment of the reagents showed different efficacies depending on BRAFV600E status. As the ratio of ERα over ERβ decreased in human thyroid cell lines which harbor wild type BRAF gene (Nthy/WT) under presence of E2, DPN, ERβ-selective agonist was more effective that MPP in Nthy/WT while Nthy/BRAFV600E (Nthy/V600E) showed the opposite. NS, not significant. aP<0.05; bP<0.01.
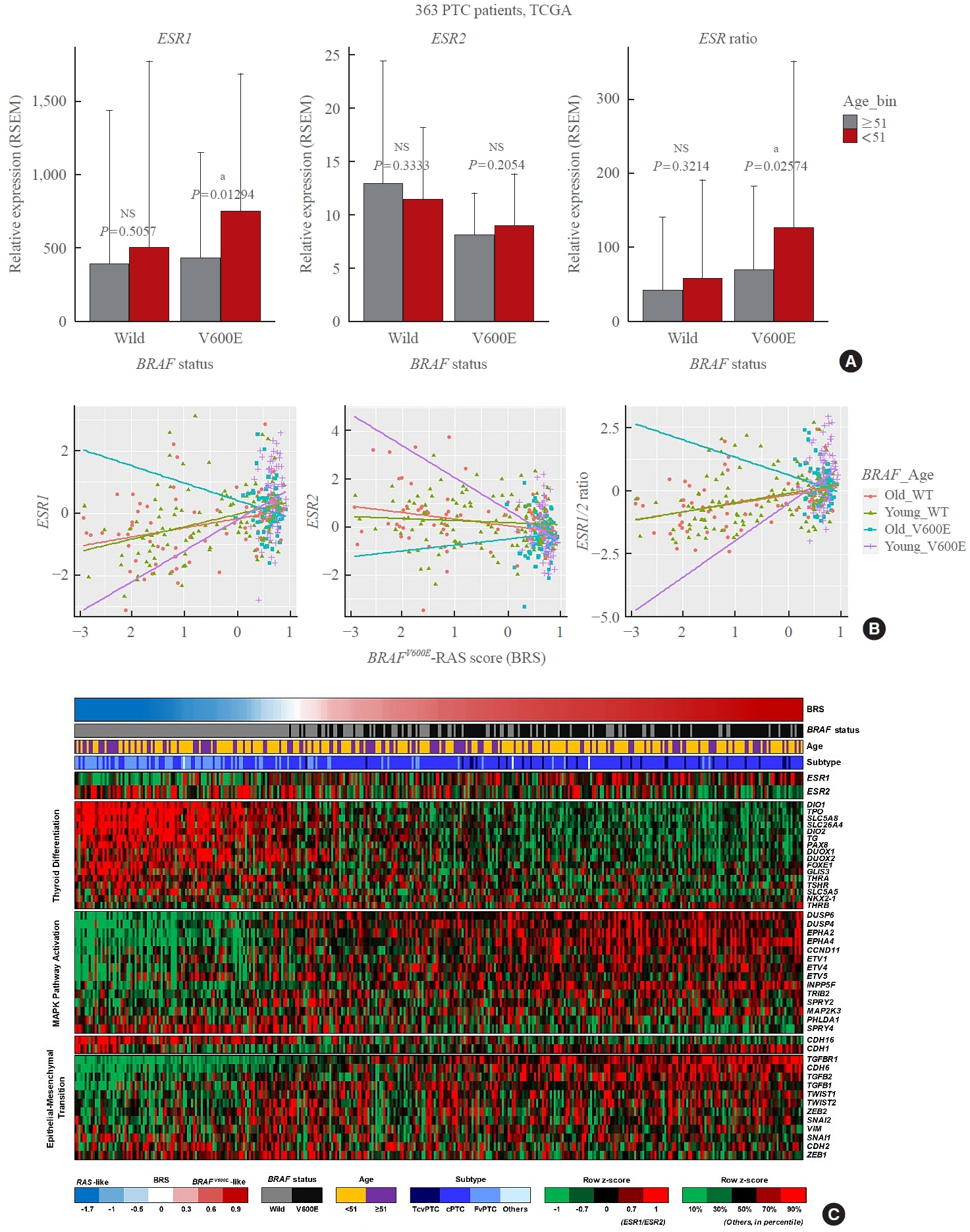
Fig. 4. Relative estrogen receptor 1 (ESR1) and ESR2 expression depending on BRAFV600E status and menopause-related age group. (A) Papillary thyroid cancer (PTC) patients (363 females) were divided according to BRAFV600E status, and we compared expression of ESR genes between age groups because the level of estrogen is known to dramatically decrease around age 50. Whereas BRAFWT group does no show significant differences in both ESR1 and ESR2 genes between age groups, BRAFV600E group shows significantly higher ESR1 and ESR1/2 ratio in younger group (age under 51). (B) Correlation between ESR genes and BRAFV600E-RAS score (BRS) in each group by BRAFV600E and age. In the BRAFWT group, there are no clear differences between the age groups, but the BRAFV600E group shows different associations depending on age group. BRAFV600E-like gene signature (higher BRS) is correlated positively with ESR1 and ESR1/2 ratio and negatively with ESR2 expression in premenopausal age group (age <51, purple line) whereas it was opposite in post-menopausal age group with BRAFV600E (age ≥51, blue line). (C) Heatmap to show ESR genes and other prognosis-related gene signatures according to the level of BRAFV600E-RAS signature in thyroid cancer. TCGA, The Cancer Genome Atlas; RSEM, RNA-sequencing by expectation maximization; NS, not significant. aP<0.05; bP<0.01.
Reference
-
1. National Cancer Institute. SEER Cancer Stat Facts: Thyroid Cancer [Internet]. Bethesda: NIH;2017. [cited 2022 Nov 28]. Available from: https://seer.cancer.gov/statfacts/html/thyro.html.2. Cancer Research UK. Thyroid cancer incidence statistics [Internet]. London: Cancer Research UK;2022. [cited 2022 Nov 28]. Available from: https://www.cancerresearchuk.org/health-professional/cancer-statistics/statistics-by-cancertype/thyroid-cancer/incidence#heading-Two.3. Horn-Ross PL, Canchola AJ, Ma H, Reynolds P, Bernstein L. Hormonal factors and the risk of papillary thyroid cancer in the California Teachers Study cohort. Cancer Epidemiol Biomarkers Prev. 2011; 20:1751–9.
Article4. Li N, Du XL, Reitzel LR, Xu L, Sturgis EM. Impact of enhanced detection on the increase in thyroid cancer incidence in the United States: review of incidence trends by socioeconomic status within the surveillance, epidemiology, and end results registry, 1980-2008. Thyroid. 2013; 23:103–10.
Article5. Yasmeen S, Cress R, Romano PS, Xing G, Berger-Chen S, Danielsen B, et al. Thyroid cancer in pregnancy. Int J Gynaecol Obstet. 2005; 91:15–20.
Article6. Gold EB. The timing of the age at which natural menopause occurs. Obstet Gynecol Clin North Am. 2011; 38:425–40.
Article7. Santoro N, Brown JR, Adel T, Skurnick JH. Characterization of reproductive hormonal dynamics in the perimenopause. J Clin Endocrinol Metab. 1996; 81:1495–501.
Article8. Rahbari R, Zhang L, Kebebew E. Thyroid cancer gender disparity. Future Oncol. 2010; 6:1771–9.
Article9. Derwahl M, Nicula D. Estrogen and its role in thyroid cancer. Endocr Relat Cancer. 2014; 21:T273–83.
Article10. Dong W, Zhang H, Li J, Guan H, He L, Wang Z, et al. Estrogen induces metastatic potential of papillary thyroid cancer cells through estrogen receptor α and β. Int J Endocrinol. 2013; 2013:941568.11. Kumar A, Klinge CM, Goldstein RE. Estradiol-induced proliferation of papillary and follicular thyroid cancer cells is mediated by estrogen receptors alpha and beta. Int J Oncol. 2010; 36:1067–80.12. Park SH, Cheung LW, Wong AS, Leung PC. Estrogen regulates Snail and Slug in the down-regulation of E-cadherin and induces metastatic potential of ovarian cancer cells through estrogen receptor alpha. Mol Endocrinol. 2008; 22:2085–98.13. Paterni I, Granchi C, Katzenellenbogen JA, Minutolo F. Estrogen receptors alpha (ERα) and beta (ERβ): subtype-selective ligands and clinical potential. Steroids. 2014; 90:13–29.
Article14. Sun Y, Wang Y, Fan C, Gao P, Wang X, Wei G, et al. Estrogen promotes stemness and invasiveness of ER-positive breast cancer cells through Gli1 activation. Mol Cancer. 2014; 13:137.
Article15. Dey P, Barros RP, Warner M, Strom A, Gustafsson JA. Insight into the mechanisms of action of estrogen receptor β in the breast, prostate, colon, and CNS. J Mol Endocrinol. 2013; 51:T61–74.
Article16. Yasar P, Ayaz G, User SD, Gupur G, Muyan M. Molecular mechanism of estrogen-estrogen receptor signaling. Reprod Med Biol. 2016; 16:4–20.
Article17. Huang Y, Dong W, Li J, Zhang H, Shan Z, Teng W. Differential expression patterns and clinical significance of estrogen receptor-α and β in papillary thyroid carcinoma. BMC Cancer. 2014; 14:383.
Article18. Yi JW, Kim SJ, Kim JK, Seong CY, Yu HW, Chai YJ, et al. Upregulation of the ESR1 gene and ESR ratio (ESR1/ESR2) is associated with a worse prognosis in papillary thyroid carcinoma: the impact of the estrogen receptor α/β expression on clinical outcomes in papillary thyroid carcinoma patients. Ann Surg Oncol. 2017; 24:3754–62.
Article19. Rajoria S, Suriano R, Shanmugam A, Wilson YL, Schantz SP, Geliebter J, et al. Metastatic phenotype is regulated by estrogen in thyroid cells. Thyroid. 2010; 20:33–41.
Article20. Xing M, Alzahrani AS, Carson KA, Viola D, Elisei R, Bendlova B, et al. Association between BRAFV600E mutation and mortality in patients with papillary thyroid cancer. JAMA. 2013; 309:1493–501.
Article21. Liu C, Chen T, Liu Z. Associations between BRAF(V600E) and prognostic factors and poor outcomes in papillary thyroid carcinoma: a meta-analysis. World J Surg Oncol. 2016; 14:241.
Article22. Lee SE, Hwang TS, Choi YL, Kim WY, Han HS, Lim SD, et al. Molecular profiling of papillary thyroid carcinoma in Korea with a high prevalence of BRAFV600E mutation. Thyroid. 2017; 27:802–10.
Article23. Atanaskova N, Keshamouni VG, Krueger JS, Schwartz JA, Miller F, Reddy KB. MAP kinase/estrogen receptor crosstalk enhances estrogen-mediated signaling and tumor growth but does not confer tamoxifen resistance. Oncogene. 2002; 21:4000–8.
Article24. Yu L, Moore AB, Castro L, Gao X, Huynh HL, Klippel M, et al. Estrogen regulates MAPK-related genes through genomic and nongenomic interactions between IGF-I receptor tyrosine kinase and estrogen receptor-alpha signaling pathways in human uterine leiomyoma cells. J Signal Transduct. 2012; 2012:204236.
Article25. Thomas RS, Sarwar N, Phoenix F, Coombes RC, Ali S. Phosphorylation at serines 104 and 106 by Erk1/2 MAPK is important for estrogen receptor-alpha activity. J Mol Endocrinol. 2008; 40:173–84.26. Marzagalli M, Montagnani Marelli M, Casati L, Fontana F, Moretti RM, Limonta P. Estrogen receptor β in melanoma: from molecular insights to potential clinical utility. Front Endocrinol (Lausanne). 2016; 7:140.
Article27. Topi G, Ghatak S, Satapathy SR, Ehrnstrom R, Lydrup ML, Sjolander A. Combined estrogen alpha and beta receptor expression has a prognostic significance for colorectal cancer patients. Front Med (Lausanne). 2022; 9:739620.
Article28. Kim BA, Jee HG, Yi JW, Kim SJ, Chai YJ, Choi JY, et al. Expression profiling of a human thyroid cell line stably expressing the BRAFV600E mutation. Cancer Genomics Proteomics. 2017; 14:53–67.
Article29. Kim M, Kim SJ, Xu Z, Ha SY, Byeon JH, Kang EJ, et al. BRAFV600E transduction of an SV40-immortalized normal human thyroid cell line induces dedifferentiated thyroid carcinogenesis in a mouse xenograft model. Thyroid. 2020; 30:487–500.
Article30. Marzagalli M, Casati L, Moretti RM, Montagnani Marelli M, Limonta P. Estrogen receptor β agonists differentially affect the growth of human melanoma cell lines. PLoS One. 2015; 10:e0134396.
Article31. Song P, Li Y, Dong Y, Liang Y, Qu H, Qi D, et al. Estrogen receptor β inhibits breast cancer cells migration and invasion through CLDN6-mediated autophagy. J Exp Clin Cancer Res. 2019; 38:354.
Article32. Zhou HB, Carlson KE, Stossi F, Katzenellenbogen BS, Katzenellenbogen JA. Analogs of methyl-piperidinopyrazole (MPP): antiestrogens with estrogen receptor alpha selective activity. Bioorg Med Chem Lett. 2009; 19:108–10.33. Cancer Genome Atlas Research Network. Integrated genomic characterization of papillary thyroid carcinoma. Cell. 2014; 159:676–90.34. Mori S, Chang JT, Andrechek ER, Matsumura N, Baba T, Yao G, et al. Anchorage-independent cell growth signature identifies tumors with metastatic potential. Oncogene. 2009; 28:2796–805.
Article35. Kim YN, Koo KH, Sung JY, Yun UJ, Kim H. Anoikis resistance: an essential prerequisite for tumor metastasis. Int J Cell Biol. 2012; 2012:306879.
Article36. Santin AP, Furlanetto TW. Role of estrogen in thyroid function and growth regulation. J Thyroid Res. 2011; 2011:875125.
Article37. Crispo F, Notarangelo T, Pietrafesa M, Lettini G, Storto G, Sgambato A, et al. BRAF inhibitors in thyroid cancer: clinical impact, mechanisms of resistance and future perspectives. Cancers (Basel). 2019; 11:1388.
Article38. Kebebew E, Weng J, Bauer J, Ranvier G, Clark OH, Duh QY, et al. The prevalence and prognostic value of BRAF mutation in thyroid cancer. Ann Surg. 2007; 246:466–71.
Article39. Weber CJ, Marvin M, Krekun S, Koschitzky T, Karp F, Benson M, et al. Effects of tamoxifen and somatostatin analogue on growth of human medullary, follicular, and papillary thyroid carcinoma cell lines: tissue culture and nude mouse xenograft studies. Surgery. 1990; 108:1065–71.40. Hoelting T, Duh QY, Clark OH, Herfarth C. Tamoxifen antagonizes proliferation and invasion of estrogen receptornegative metastatic follicular thyroid cancer cells via protein kinase C. Cancer Lett. 1996; 100:89–93.
Article41. de Araujo LF, Soares JM Jr, Simoes RS, Calio PL, OliveiraFilho RM, Simoes Mde J, et al. Effect of conjugated equine estrogens and tamoxifen administration on thyroid gland histomorphology of the rat. Clinics (Sao Paulo). 2006; 61:321–6.
Article42. Zidan J, Rubenstein W. Effect of adjuvant tamoxifen therapy on thyroid function in postmenopausal women with breast cancer. Oncology. 1999; 56:43–5.
Article43. Jiang D, Srinivasan A, Lozano G, Robbins PD. SV40 T antigen abrogates p53-mediated transcriptional activity. Oncogene. 1993; 8:2805–12.44. DeCaprio JA, Ludlow JW, Figge J, Shew JY, Huang CM, Lee WH, et al. SV40 large tumor antigen forms a specific complex with the product of the retinoblastoma susceptibility gene. Cell. 1988; 54:275–83.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Clinical Implication of BRAF Mutation in Thyroid Cancer
- Expressions of miRNAs in Papillary Thyroid Carcinoma and Their Associations with the BRAFV600EMutation and Clinicopathological Features.
- Association of BRAF(V600E) Mutation with Poor Clinical Prognostic Factors and Ultrasonographic Findings in Cases of Papillary Thyroid Carcinoma
- Clinicopathological Implications of the BRAF(V600E) Mutation in PTC with Concurrent Hashimoto Thyroiditis
- Immunocytochemical analysis for estrogen receptors in the patients with thyroid disease
