Neuroprotective Effects of GV1001 in Animal Stroke Model and Neural Cells Subject to Oxygen-Glucose Deprivation/Reperfusion Injury
- Affiliations
-
- 1Department of Neurology, Hanyang University Guri Hospital, Hangyang University College of Medicine, Guri, Korea
- 2Department of Neurology, Hanyang University Seoul Hospital, Hanyang University College of Medicine, Seoul, Korea
- 3Department of Translational Medicine, Hanyang University Graduate School of Biomedical Science & Engineering, Seoul, Korea
- KMID: 2520919
- DOI: http://doi.org/10.5853/jos.2021.00626
Abstract
- Background and Purpose
Previous studies have revealed the diverse neuroprotective effects of GV1001. In this study, we investigated the effects of GV1001 on focal cerebral ischemia-reperfusion injury (IRI) in rats and oxygen-glucose deprivation/reoxygenation (OGD/R)-induced injury in neural stem cells (NSCs) and cortical neurons.
Methods
Focal cerebral IRI was induced by transient middle cerebral artery occlusion (MCAO). Brain diffusion-weighted imaging (DWI) was performed 2 hours after occlusion, and a total of 37 rats were treated by reperfusion with GV1001 or saline 2 hours after occlusion. Fluid-attenuated inversion recovery (FLAIR) magnetic resonance imaging, immunohistochemistry, and neurobehavioral function analyses were performed. Additionally, OGD/R-injured NSCs and cortical neurons were treated with different GV1001 concentrations. Cell viability, proliferation, migration, and oxidative stress were determined by diverse molecular analyses.
Results
In the stroke model, GV1001 protected neural cells against IRI. The most effective dose of GV1001 was 60 μM/kg. The infarct volume on FLAIR 48 hours after MCAO compared to lesion volume on DWI showed a significantly smaller ratio in the GV1001-treated group. GV1001-treated rats exhibited better behavioral functions than the saline-treated rats. Treatment with GV1001 increased the viability, proliferation, and migration of the OGD/R-injured NSCs. Free radicals were significantly restored by treatment with GV1001. These neuroprotective effects of GV1001 have also been demonstrated in OGD/R-injured cortical neurons. Conclusions The results suggest that GV1001 has neuroprotective effects against IRI in NSCs, cortical neurons, and the rat brain. These effects are mediated through the induction of cellular proliferation, mitochondrial stabilization, and anti-apoptotic, anti-aging, and antioxidant effects.
Figure
-
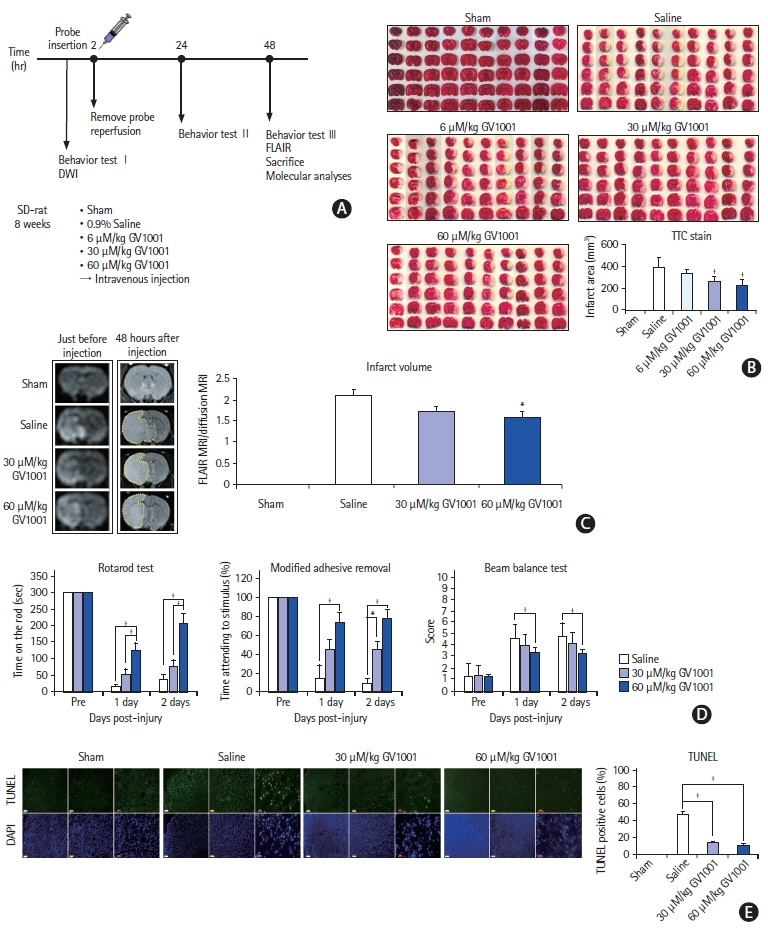
Figure 1. (A) Schematic representation of the study design and the timeline for the establishment of the ischemic stroke animal model. (B) 2,3,5-Triphenyltetrazolium chloride staining showing the amount of cerebral infarction in each group. The infarct volume was significantly reduced in groups treated with 30 and 60 μM/kg GV1001. (C) The infarct volume was measured by diffusion-weighted imaging (DWI) and fluid-attenuated inversion recovery (FLAIR) magnetic resonance imaging (MRI). The volume was significantly reduced in the 60 μM/kg GV1001 group. (D) All behavioral function tests showed a beneficial effect of GV1001. (E) The number of transferase-mediated deoxyuridine triphosphate nick end labeling (TUNEL)-positive cells was reduced in the GV1001-treated group, white scale bar: 250 μm; yellow scale bar: 100 μm; and red scale bar: 25 μm. SD, Sprague-Dawley; TTC, triphenyltetrazolium chloride; DAPI, 4',6-diamidino-2-phenylindole. *P<0.05; †P<0.01 (vs. control-saline group).
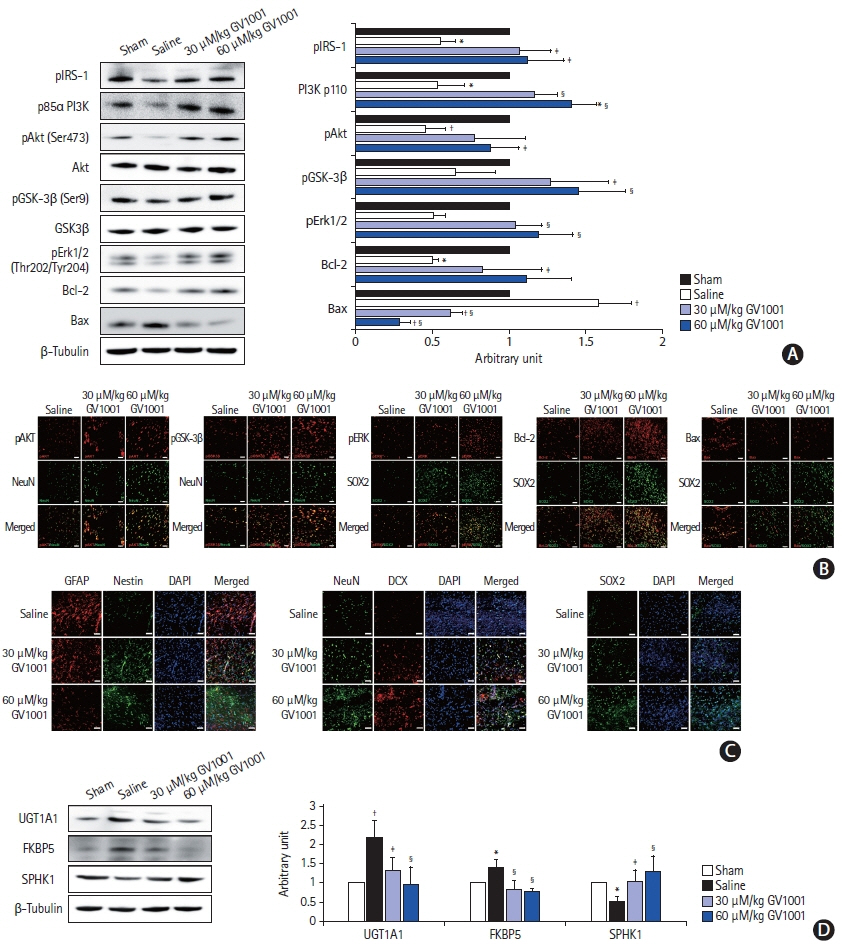
Figure 2. The effect of GV1001 on intracellular signaling proteins and cells. (A) Decreased levels of phosphorylated Akt (pAkt; Ser473), phospho-glycogen synthase kinase (pGSK-3β; Ser9), phosphorylated-extracellular signal-regulated kinase (pERK), B-cell lymphoma 2 (Bcl-2), and increased levels of Bcl-2 associated X (Bax) were noted in the peri-infarct regions of GV1001-treated rats compared to those of control rats. Administration of GV1001 (30 and 60 μM/kg) alleviated the changes. (B) Immunohistochemistry (IHC) staining showing increased pAkt (Ser473), pGSK-3β (Ser9), pERK, Bcl-2 levels and decreased Bax levels along with increased numbers of neuronal nuclei (NeuN)- or SRY-box transcription factor 2 (SOX2)-positive cells in GV1001-treated groups, scale bar: 50 μm. (C) IHC also revealed increased levels of nestin (a neuroectodermal stem cell marker), NeuN (a neuronal nuclear antigen), doublecortin (DCX; a neuronal differentiation marker), and SOX2 (a multipotent neural stem cell marker) in the peri-infarct region of GV1001-treated rats. Glial fibrillary acidic protein (GFAP; an astrocyte marker) expression was decreased in GV1001-treated rats, scale bar: 50 μm. (D) Levels of neurotoxic phenotype reactive astrocytes (uridine diphosphate glucuronosyltransferase 1A1 [UGT1A1] and FK506 binding protein 5 [FKBP5]) were increased and those of neuroprotective phenotype reactive astrocytes (sphingosine kinase type 1 [SPHK1]) were decreased in the peri-infarct area, but GV1001 significantly restored the expression of these markers. pIRS-1, phospho-insulin receptor substrate-1; DAPI, 4',6-diamidino-2-phenylindole. *P<0.05 (vs. sham group); †P<0.01 (vs. sham group); ‡P<0.05 (vs. saline group); §P<0.01 (vs. saline group).
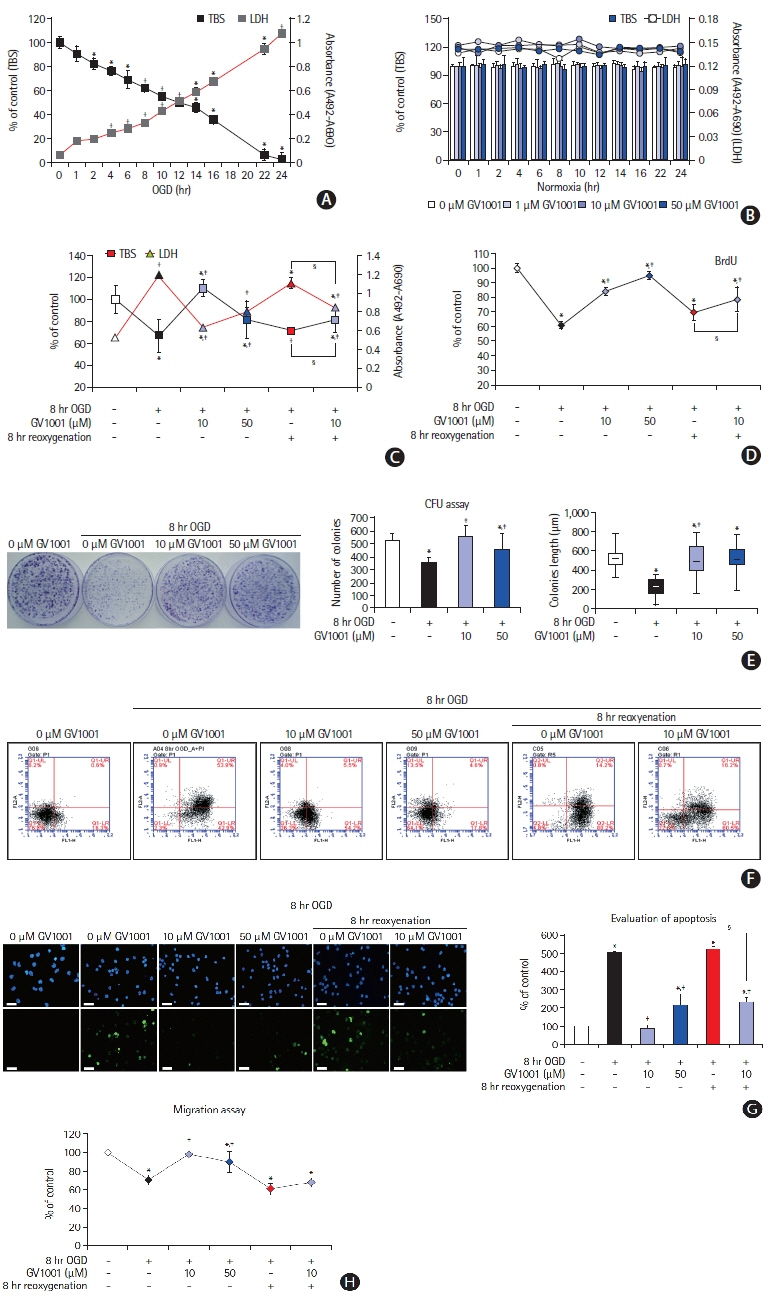
Figure 3. Effect of GV1001 on viability, cytotoxicity, and proliferation of neural stem cells (NSCs) after treatment with oxygen-glucose deprivation (OGD) alone or OGD/reperfusion (OGD/R). (A) OGD induced cell death and cytotoxicity in a time-dependent manner. (B) GV1001 at concentrations of up to 50 μM did not affect the viability and cytotoxicity of NSCs. (C) Treatment with GV1001 increased cell viability and decreased cytotoxicity of NSCs injured by OGD/R. (D, E) To evaluate the effects of prolonged OGD/R and/or varying concentrations of GV1001 on cell proliferation, NSCs were exposed to OGD/R for 8 hours with different concentrations of GV1001 and were incubated in bromodeoxyuridine (BrdU) labeling medium. CFU assays showed that proliferation was inhibited in OGD/R-injured NSCs but was markedly restored with GV1001 treatment. (F) Various viability tests also showed that GV1001 increased the viability of NSCs after injury by OGD/R. (G) 4',6-Diamidino-2-phenylindole (DAPI) and transferasemediated deoxyuridine triphosphate nick end labeling (TUNEL) staining revealed the anti-apoptotic effect of GV1001 on OGD/R-injured NSCs, scale bar: 25 μm. (H) Cell migration assay showed that treatment with GV1001 restored cell migration activity of NSCs that had been reduced by OGD/R. TBS, trypan blue staining; LDH, lactate dehydrogenase; CFU, colony-forming unit. *P<0.05 (vs. control group); †P<0.01 (vs. control group); ‡P<0.05 (vs. NSCs only treated with OGD); §P<0.05 (vs. NSCs treated with OGD/R).

Figure 4. (A) Antioxidant effect of GV1001. GV1001 alone did not trigger reactive oxygen species (ROS) production. (B) Intracellular ROS produced after oxygenglucose deprivation/reperfusion (OGD/R) was decreased after treatment with GV1001, scale bar: 25 μm. (C, D, E) The levels of malondialdehyde (MDA) and intracellular Ca2+ and oxidative mitochondrial DNA damage were increased by OGD/R and were significantly decreased upon GV1001 treatment. (F) Adenosine triphosphate (ATP) assay for the function of mitochondria. ATP levels were decreased by OGD/R. GV1001 restored ATP levels. (G) The mitochondrial membrane potential was significantly decreased with OGD/R, but GV1001 markedly restored this potential. mtDNA, mitochondrial deoxyribonucleic acid. *P<0.05 (vs. control group); †P<0.01 (vs. control group); ‡P<0.05 (vs. NSCs only treated with OGD); §P<0.01 (vs. NSCs only treated with OGD); ∥P<0.05 (vs. NSCs treated with OGD/R).

Figure 5. (A) Effect of GV1001 on intracellular proteins in oxygen-glucose deprivation/reoxygenation (OGD/R)-injured neural stem cells (NSCs). Western blots showed that the levels of proteins associated with the proliferation and survival of NSCs decreased with OGD/R. The protein levels were increased upon treatment with GV1001. (B) Levels of proteins associated with cell death increased with OGD/R. These levels were decreased upon GV1001 treatment. (C) Expressions of active β-catenin, cyclin D1, and C-myc were evaluated to confirm the non-canonical telomerase activity of GV1001. These levels were decreased after OGD/R and were restored by GV1001 treatment. The data are expressed as percentages of the value in the control group and represented as the mean±standard deviation values from five independent experiments. (D) Proteomic and (E) antibody microarray data revealed that GV1001 affected the levels of numerous intracellular proteins in OGD/R-injured NSCs. PI3K, phosphoinositide 3-kinase; pAkt, phosphorylated Akt; pGSK-3β, phospho-glycogen synthase kinase; HMGB1, high mobility group box protein 1; Bax, Bcl-2 associated X; HSP, heat shock protein; ATP, adenosine triphosphate; PP1, protein phosphatase 1; NADH, nicotinamide adenine dinucleotide hydride; COP9, constitutive photomorphogenesis 9; CoA, coenzyme A; GRP1, general receptor of phosphoinositides 1; ARNO3; ARF nucleotide-binding site opener 3; ERK, extracellular signal-regulated kinase; b-NOS, brain nitric oxide synthase; PKB, protein kinase B; PKC, protein kinase C. *P<0.05 (vs. control group); †P<0.01 (vs. control group); ‡P<0.05 (vs. NSCs only treated with OGD); §P<0.01 (vs. NSCs only treated with OGD); ∥P<0.05 (vs. NSCs treated with OGD/R).
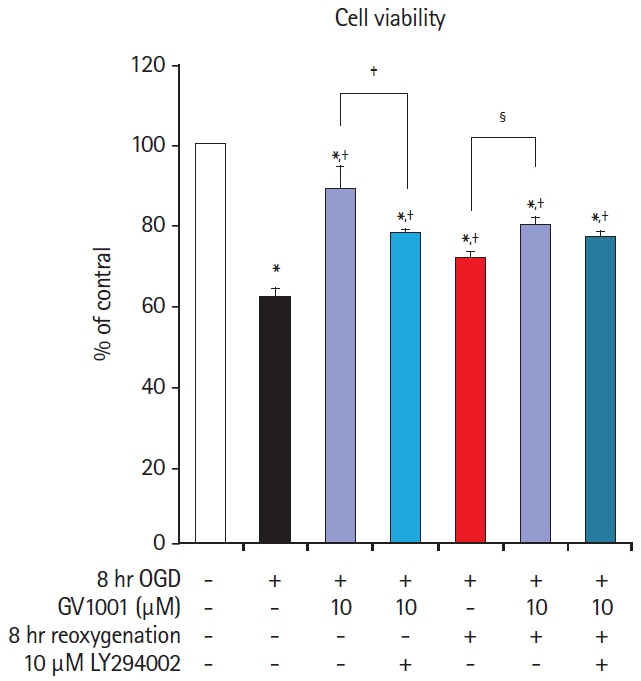
Figure 6. Oxygen-glucose deprivation/reoxygenation (OGD/R)-injured neural stem cells (NSCs) were treated with the phosphoinositide 3-kinase (PI3K) inhibitor LY294002 to assess the role of the PI3K/Akt pathway in the protective mechanism of GV1001. The NSCs were divided into seven groups: control (group 1); OGD (group 2); OGD+10 μM GV1001 (group 3); OGD+10 μM GV1001+LY294002 (group 4); OGD/R (group 5); OGD/R+10 μM GV1001 (group 6); and OGD/R+10 μM GV1001+LY294002 (group 7). LY294002 significantly blocked the effects of GV1001 on OGD-injured NSCs. *P<0.05 (vs. group 1); †P<0.05 (vs. group 2); ‡P<0.05 (group 3 vs. group 4); §P<0.05 (group 5 vs. group 6).
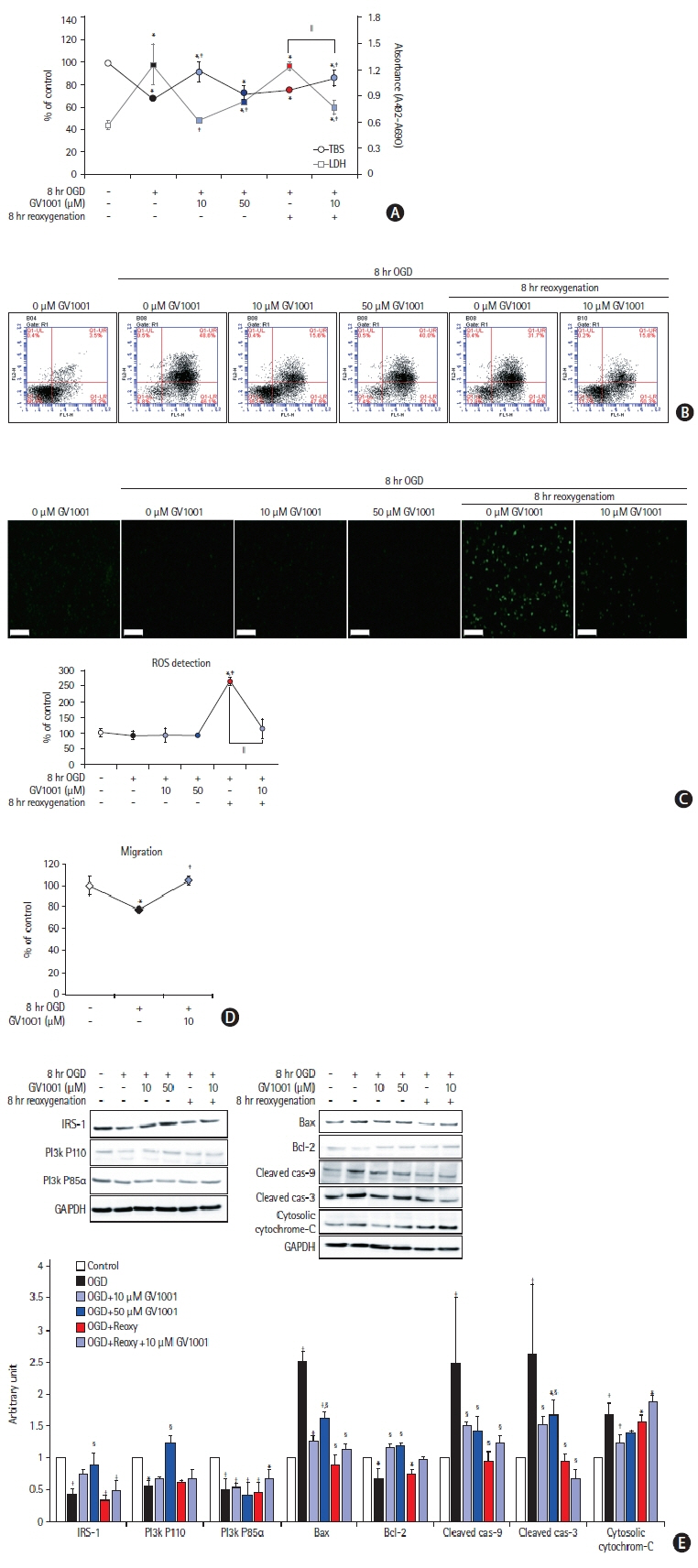
Figure 7. Effects of GV1001 on cortical neurons injured by oxygen-glucose deprivation/reoxygenation (OGD/R). GV1001 restored the viability (A) and alleviated the toxicity (B) of cortical neurons injured by OGD/R. (C) GV1001 significantly inhibited the production of reactive oxygen species (ROS), which was elevated by reoxygenation after OGD, in neurons, scale bar: 100 μm. (D) Treatment with GV1001 restored the migration activity of cortical neurons, which was reduced by OGD/R. (E) OGD/R decreased the levels of survival-related proteins and increased those of death-associated proteins. Treatment with GV1001 reversed these changes. TBS, trypan blue staining; LDH, lactate dehydrogenase; IRS-1, insulin receptor substrate; PI3K, phosphoinositide 3-kinase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; Bax, Bcl-2 associated X. *P<0.05 (vs. control group); †P<0.01 (vs. control group); ‡P<0.05 (vs. NSCs only treated with OGD); §P<0.01 (vs. NSCs only treated with OGD); ∥P<0.05 (vs. NSCs treated with OGD/R).
Reference
-
References
1. Vidale S, Consoli A, Arnaboldi M, Consoli D. Postischemic Inflammation in Acute Stroke. J Clin Neurol. 2017; 13:1–9.
Article2. Jung JE, Kim GS, Chen H, Maier CM, Narasimhan P, Song YS, et al. Reperfusion and neurovascular dysfunction in stroke: from basic mechanisms to potential strategies for neuroprotection. Mol Neurobiol. 2010; 41:172–179.
Article3. Park HH, Yu HJ, Kim S, Kim G, Choi NY, Lee EH, et al. Neural stem cells injured by oxidative stress can be rejuvenated by GV1001, a novel peptide, through scavenging free radicals and enhancing survival signals. Neurotoxicology. 2016; 55:131–141.
Article4. Martínez P, Blasco MA. Telomeric and extra-telomeric roles for telomerase and the telomere-binding proteins. Nat Rev Cancer. 2011; 11:161–176.
Article5. Park HH, Lee KY, Kim S, Lee JW, Choi NY, Lee EH, et al. Novel vaccine peptide GV1001 effectively blocks β-amyloid toxicity by mimicking the extra-telomeric functions of human telomerase reverse transcriptase. Neurobiol Aging. 2014; 35:1255–1274.6. Ko YJ, Kwon KY, Kum KY, Lee WC, Baek SH, Kang MK, et al. The anti-inflammatory effect of human telomerase-derived peptide on P. gingivalis lipopolysaccharide-induced inflammatory cytokine production and its mechanism in human dental pulp cells. Mediators Inflamm. 2015; 2015:385127.7. Koh SH, Park Y, Song CW, Kim JG, Kim K, Kim J, et al. The effect of PARP inhibitor on ischaemic cell death, its related inflammation and survival signals. Eur J Neurosci. 2004; 20:1461–1472.
Article8. Koh SH, Yoo AR, Chang DI, Hwang SJ, Kim SH. Inhibition of GSK-3 reduces infarct volume and improves neurobehavioral functions. Biochem Biophys Res Commun. 2008; 371:894–899.
Article9. Kim YS, Yoo A, Son JW, Kim HY, Lee YJ, Hwang S, et al. Early activation of phosphatidylinositol 3-kinase after ischemic stroke reduces infarct volume and improves long-term behavior. Mol Neurobiol. 2017; 54:5375–5384.
Article10. Chen J, Li Y, Wang L, Zhang Z, Lu D, Lu M, et al. Therapeutic benefit of intravenous administration of bone marrow stromal cells after cerebral ischemia in rats. Stroke. 2001; 32:1005–1011.
Article11. Sughrue ME, Mocco J, Komotar RJ, Mehra A, D’Ambrosio AL, Grobelny BT, et al. An improved test of neurological dysfunction following transient focal cerebral ischemia in rats. J Neurosci Methods. 2006; 151:83–89.
Article12. Studer L, Tabar V, McKay RD. Transplantation of expanded mesencephalic precursors leads to recovery in parkinsonian rats. Nat Neurosci. 1998; 1:290–295.
Article13. Currle DS, Hu JS, Kolski-Andreaco A, Monuki ES. Culture of mouse neural stem cell precursors. J Vis Exp. 2007; 2:152.
Article14. Koo TY, Yan JJ, Yang J. Protective effect of peptide GV1001 against renal ischemia-reperfusion injury in mice. Transplant Proc. 2014; 46:1117–1122.
Article15. Kyte JA. Cancer vaccination with telomerase peptide GV1001. Expert Opin Investig Drugs. 2009; 18:687–694.
Article16. Lee YK, Nata’atmaja BS, Kim BH, Pak CS, Heo CY. Protective effect of telomerase-based 16-mer peptide vaccine (GV1001) on inferior epigastric island skin flap survivability in ischaemia-reperfusion injury rat model. J Plast Surg Hand Surg. 2017; 51:210–216.
Article17. Chang JE, Kim HJ, Jheon S, Lim C. Protective effects of GV1001 on myocardial ischemia-reperfusion injury. Mol Med Rep. 2017; 16:7315–7320.
Article18. Chang JE, Kim HJ, Yi E, Jheon S, Kim K. Reduction of ischaemiareperfusion injury in a rat lung transplantation model by low-concentration GV1001. Eur J Cardiothorac Surg. 2016; 50:972–979.
Article19. Son JW, Choi H, Yoo A, Park HH, Kim YS, Lee KY, et al. Activation of the phosphatidylinositol 3-kinase pathway plays important roles in reduction of cerebral infarction by cilnidipine. J Neurochem. 2015; 135:186–193.
Article20. Pap M, Cooper GM. Role of translation initiation factor 2B in control of cell survival by the phosphatidylinositol 3-kinase/Akt/glycogen synthase kinase 3beta signaling pathway. Mol Cell Biol. 2002; 22:578–586.21. Hemmings BA, Restuccia DF. The PI3K-PKB/Akt pathway. Cold Spring Harb Perspect Biol. 2015; 7:a026609.
Article22. Song WJ, Song EA, Jung MS, Choi SH, Baik HH, Jin BK, et al. Phosphorylation and inactivation of glycogen synthase kinase 3β (GSK3β) by dual-specificity tyrosine phosphorylation-regulated kinase 1A (Dyrk1A). J Biol Chem. 2015; 290:2321–2333.
Article23. Yamaguchi H, Wang HG. The protein kinase PKB/Akt regulates cell survival and apoptosis by inhibiting Bax conformational change. Oncogene. 2001; 20:7779–7786.
Article24. Xie C, Zhou M, Lin J, Wu Z, Ding S, Luo J, et al. EEF1D promotes glioma proliferation, migration, and invasion through EMT and PI3K/Akt pathway. Biomed Res Int. 2020; 2020:7804706.
Article25. Hemmings BA, Restuccia DF. PI3K-PKB/Akt pathway. Cold Spring Harb Perspect Biol. 2012; 4:a011189.
Article26. Quan C, Sun J, Lin Z, Jin T, Dong B, Meng Z, et al. Ezrin promotes pancreatic cancer cell proliferation and invasion through activating the Akt/mTOR pathway and inducing YAP translocation. Cancer Manag Res. 2019; 11:6553–6566.27. Koh SH, Park HH. Neurogenesis in stroke recovery. Transl Stroke Res. 2017; 8:3–13.
Article28. Zhang S, Lachance BB, Moiz B, Jia X. Optimizing stem cell therapy after ischemic brain injury. J Stroke. 2020; 22:286–305.
Article29. Ellis P, Fagan BM, Magness ST, Hutton S, Taranova O, Hayashi S, et al. SOX2, a persistent marker for multipotential neural stem cells derived from embryonic stem cells, the embryo or the adult. Dev Neurosci. 2004; 26:148–165.
Article30. Wei ZZ, Zhang JY, Taylor TM, Gu X, Zhao Y, Wei L. Neuroprotective and regenerative roles of intranasal Wnt-3a administration after focal ischemic stroke in mice. J Cereb Blood Flow Metab. 2018; 38:404–421.
Article31. Richardsen E, Andersen S, Al-Saad S, Rakaee M, Nordby Y, Pedersen MI, et al. Evaluation of the proliferation marker Ki-67 in a large prostatectomy cohort. PLoS One. 2017; 12:e0186852.
Article32. Lange SS, Mitchell DL, Vasquez KM. High mobility group protein B1 enhances DNA repair and chromatin modification after DNA damage. Proc Natl Acad Sci U S A. 2008; 105:10320–10325.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The individual and combined neuroprotective effects of propofol and ketamine on rat mixed cortical cultures exposed to oxygen-glucose deprivation-reperfusion injury
- The influence of propofol administration time on oxygen-glucose deprivation-reperfusion injury in rat mixed cortical cultures focused on N-Methyl-D-Aspartate (NMDA) receptors
- BMP‑6 Attenuates Oxygen and Glucose Deprivation-Induced Apoptosis in Human Neural Stem Cells through Inhibiting p38 MAPK Signaling Pathway
- The optimal model of reperfusion injury in vitro using H9c2 transformed cardiac myoblasts
- Gene Expression Profile in Microglia following Ischemia-Reperfusion Injury
