MDL-12330A potentiates TRAIL-induced apoptosis in gastric cancer cells through CHOP-mediated DR5 upregulation
- Affiliations
-
- 1Department of Pathology, College of Medicine, Chosun University, Gwangju 61501, Korea.
- 2Division of Premedical Science, College of Medicine, Chosun University, Gwangju 61501, Korea. sihan@chosun.ac.kr
- KMID: 2384453
- DOI: http://doi.org/10.4196/kjpp.2017.21.4.397
Abstract
- MDL-12330A is a widely used adenylyl cyclase (AC) inhibitor that blocks AC/cAMP signaling. In this study, we demonstrated a novel antitumor activity of this drug in gastric carcinoma (GC) cell lines. In these GC cells, MDL-12330A reduced cell viability and induced cell death in a concentration-dependent manner. At a moderate concentration (~20 µM), MDL-12330A mainly induced apoptotic death whereas at concentrations greater than 20 µM, it increased non-apoptotic cell death. The induction of apoptosis was at least partially regulated by CHOP-mediated DR5 upregulation, as detected by immunoblotting and gene interference assays. More importantly, low concentrations of MDL-12330A effectively enhanced recombinant human tumor necrosis factor (TNF)-related apoptosis-inducing ligand (rhTRAIL)-induced apoptosis and clonogenicity in these gastric cancer cells. This study demonstrates a possible role of MDL-12330A as a potential sensitizer to TRAIL, and suggests a novel therapeutic strategy targeting gastric cancer cells.
Keyword
MeSH Terms
Figure
-
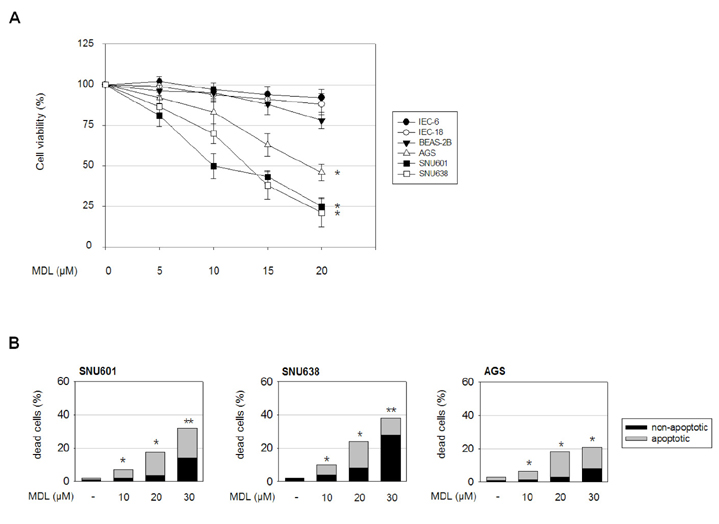
Fig. 1 MDL-12330A induces an antitumor effect in human GC cells. (A, B) Cells were treated with the indicated concentrations of MDL-12330A (MDL) for 24 h and subjected to the EZ-cytox assay for measurement of cell viability (A), or stained with Hoechst 33342/PI to assess the number of cells undergoing apoptotic or non-apoptotic cell death (B) as described in the Methods section. *p<0.05, **p<0.01 vs. control.
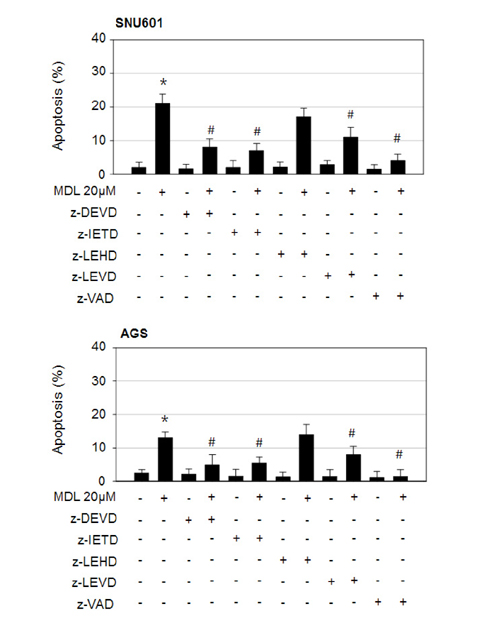
Fig. 2 Apoptosis triggered by the combined action of MDL-12330A and rhTRAIL involves activation of caspase-3, caspase-8, and caspase-4, but not caspase-9. SNU601 and AGS cells were treated with 20 µM MDL-12330A in the absence or presence of z-DEVD, z-IETD, z-LEHD, z-LEVD, or z-VAD for 24 h. The treated cells were stained with Hoechst33342, and apoptotic body was counted under a fluorescence microscope. The number of apoptotic cells was expressed as a percentage of the total number of cells counted. *p<0.05 vs. control; #p<0.05 vs. MDL-12330A (MDL) treated cells.

Fig. 3 DR5 plays an important role in MDL-12330A-mediated apoptosis. (A) SNU601 cells were exposed to 20 µM MDL-12330A, and the mRNA level of DR5 was determined using real-time PCR. (B) SNU601, SNU638, and AGS cells were treated with various concentrations of MDL-12330A for 24 h, and then analyzed by immunoblotting with antibodies against DR5, DR4, or α-tubulin. (C) SNU601 cells transfected with scrambled small interfering RNA (CTL RNAi), DR5 RNAi or DR4 RNAi were treated with 20 µM MDL-12330A for 24 h (left graph), or 10 µM MDL-12330A for 24 h (right graph). Treated cells were then stained using Hoechst 33342, and apoptotic body was counted under a fluorescence microscope. The number of apoptotic cells was expressed as a percentage of the total number of cells counted. *p<0.05 vs. control; #p<0.05 vs. MDL-12330A treated and CTL RNAi cells.

Fig. 4 MDL-12330A-mediated DR5 expression is regulated by CHOP. (A) SNU601 or SNU638 cells were transfected with a scrambled small interfering RNA (CTL RNAi), p65 RNAi, CHOP RNAi, and C-EBPβ RNAi, and then treated with 20 µM MDL-12330A (MDL) for 24 h. Cell lysates were prepared and analyzed by immunoblotting to assess DR5 expression. Silencing effect of each siRNA was confirmed by immunoblotting in vehicle treated control samples. (B, C) SNU601 or SNU638 cells were exposed to the indicated concentrations of MDL-12330A for 16 h and cell lysates were analyzed by immunoblotting with an antibody to CHOP (B), and to BiP and p-PERK (C). Alpha-tubulin was used as a loading control.
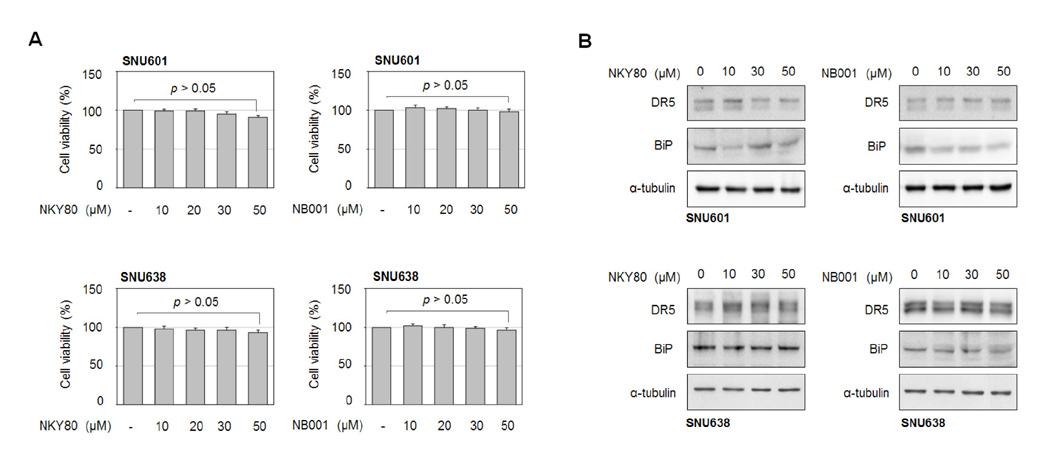
Fig. 5 Antitumor effect of MDL-12330A is not linked to inhibition of adenylate cyclase activity. (A) SNU601 and SNU638 cells were exposed to other type of adenylate cyclase inhibitors NKY80 and NB001 at indicated concentrations for 24 h, and cell viability was assessed by the EZ-cytox assay. (B) SNU601 and SNU638 cells were incubated with NKY80 or NB001 at 0, 10, 30 and 50 µM for 24 h, and cell lysates were prepared and analyzed by immunoblotting with an antibody to DR5 and BiP.
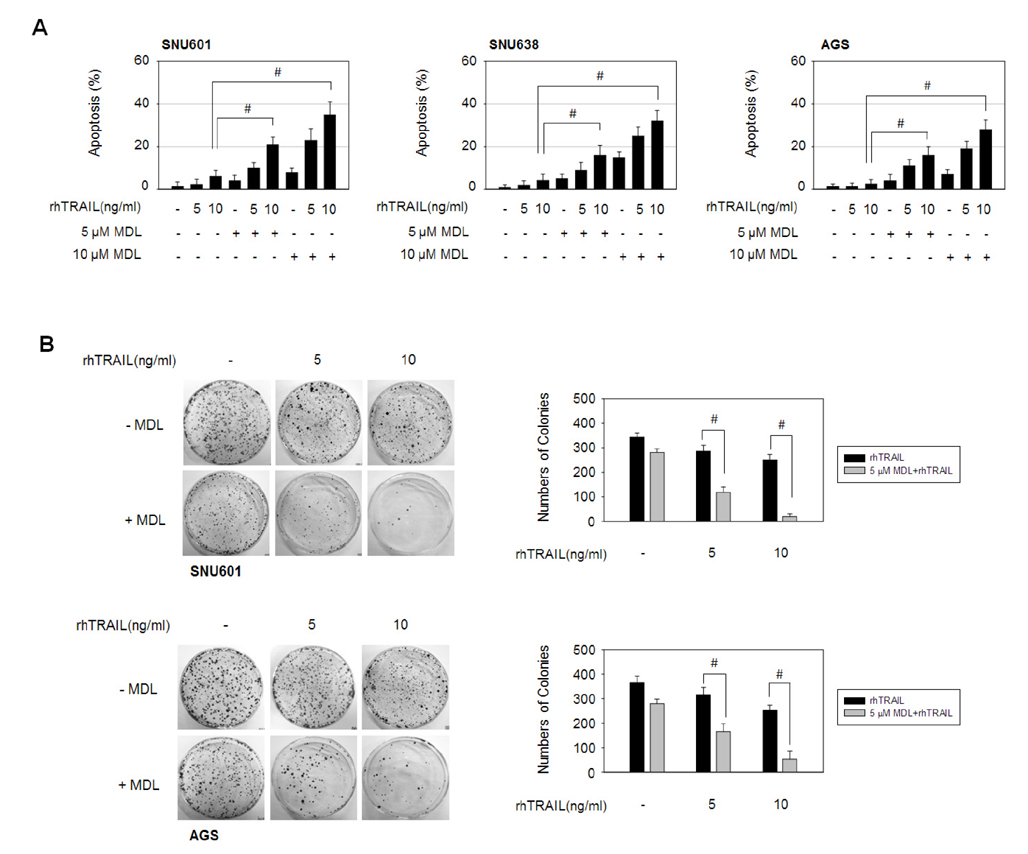
Fig. 6 MDL-12330A enhances rhTRAIL-induced apoptosis and clonogenicity. (A) Cells were pre-treated with 5 or 10 µM MDL-12330A for 24 h and then further exposed to rhTRAIL for 6 h. Apoptosis was detected by staining cells with Hoechst 33342 and assessing the ratio of apoptotic nuclei to normal nuclei under a fluorescence microscope. (B) Cells were incubated in the absence or presence of 5 µM MDL-12330A for 24 h and then exposed to 0, 5, or 10 ng/ml rhTRAIL for the last 2 h. Following this, 2×103 cells were re-plated on 60mm dishes. Colonies were stained with crystal violet and counted at 2 weeks post-incubation. #p<0.05 vs. rhTRAIL treated alone.
Reference
-
1. Pisani P, Parkin DM, Bray F, Ferlay J. Estimates of the worldwide mortality from 25 cancers in 1990. Int J Cancer. 1999; 83:18–29.2. Greenlee RT, Hill-Harmon MB, Murray T, Thun M. Cancer statistics, 2001. CA Cancer J Clin. 2001; 51:15–36.3. Roukos DH. Current status and future perspectives in gastric cancer management. Cancer Treat Rev. 2000; 26:243–255.4. Gura T. How TRAIL kills cancer cells, but not normal cells. Science. 1997; 277:768.5. Baker SJ, Reddy EP. Modulation of life and death by the TNF receptor superfamily. Oncogene. 1998; 17:3261–3270.6. Ashkenazi A. Targeting death and decoy receptors of the tumour-necrosis factor superfamily. Nat Rev Cancer. 2002; 2:420–430.7. Srivastava RK. TRAIL/Apo-2L: mechanisms and clinical applications in cancer. Neoplasia. 2001; 3:535–546.8. Prasad S, Yadav VR, Kannappan R, Aggarwal BB. Ursolic acid, a pentacyclin triterpene, potentiates TRAIL-induced apoptosis through p53-independent up-regulation of death receptors: evidence for the role of reactive oxygen species and JNK. J Biol Chem. 2011; 286:5546–5557.9. Siddiqui IA, Malik A, Adhami VM, Asim M, Hafeez BB, Sarfaraz S, Mukhtar H. Green tea polyphenol EGCG sensitizes human prostate carcinoma LNCaP cells to TRAIL-mediated apoptosis and synergistically inhibits biomarkers associated with angiogenesis and metastasis. Oncogene. 2008; 27:2055–2063.10. Szliszka E, Krol W. The role of dietary polyphenols in tumor necrosis factor-related apoptosis inducing ligand (TRAIL)-induced apoptosis for cancer chemoprevention. Eur J Cancer Prev. 2011; 20:63–69.11. Seifert R, Lushington GH, Mou TC, Gille A, Sprang SR. Inhibitors of membranous adenylyl cyclases. Trends Pharmacol Sci. 2012; 33:64–78.12. Li X, Guo Q, Gao J, Yang J, Zhang W, Liang Y, Wu D, Liu Y, Weng J, Li Q, Zhang Y. The adenylyl cyclase inhibitor MDL-12,330A potentiates insulin secretion via blockade of voltage-dependent K(+) channels in pancreatic beta cells. PLoS One. 2013; 8:e77934.13. Gadea A, López E, López-Colomé AM. The adenylate cyclase inhibitor MDL-12330A has a non-specific effect on glycine transport in Müller cells from the retina. Brain Res. 1999; 838:200–204.14. Shin JN, Park SY, Cha JH, Park JY, Lee BR, Jung SA, Lee ST, Yun CW, Seol DW, Kim TH. Generation of a novel proform of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) protein that can be reactivated by matrix metalloproteinases. Exp Cell Res. 2006; 312:3892–3898.15. Kim CH, Han SI, Lee SY, Youk HS, Moon JY, Duong HQ, Park MJ, Joo YM, Park HG, Kim YJ, Yoo MA, Lim SC, Kang HS. Protein kinase C-ERK1/2 signal pathway switches glucose depletion-induced necrosis to apoptosis by regulating superoxide dismutases and suppressing reactive oxygen species production in A549 lung cancer cells. J Cell Physiol. 2007; 211:371–385.16. Franken NA, Rodermond HM, Stap J, Haveman J, van Bree C. Clonogenic assay of cells in vitro. Nat Protoc. 2006; 1:2315–2319.17. Kong F, You H, Zhao J, Liu W, Hu L, Luo W, Hu W, Tang R, Zheng K. The enhanced expression of death receptor 5 (DR5) mediated by HBV X protein through NF-kappaB pathway is associated with cell apoptosis induced by (TNF-α related apoptosis inducing ligand) TRAIL in hepatoma cells. Virol J. 2015; 12:192.18. Wu GS, Burns TF, McDonald ER 3rd, Jiang W, Meng R, Krantz ID, Kao G, Gan DD, Zhou JY, Muschel R, Hamilton SR, Spinner NB, Markowitz S, Wu G, el-Deiry WS. KILLER/DR5 is a DNA damage-inducible p53-regulated death receptor gene. Nat Genet. 1997; 17:141–143.19. Pennati M, Sbarra S, De Cesare M, Lopergolo A, Locatelli SL, Campi E, Daidone MG, Carlo-Stella C, Gianni AM, Zaffaroni N. YM155 sensitizes triple-negative breast cancer to membrane-bound TRAIL through p38 MAPK- and CHOP-mediated DR5 upregulation. Int J Cancer. 2015; 136:299–309.20. Lim JH, Park JW, Choi KS, Park YB, Kwon TK. Rottlerin induces apoptosis via death receptor 5 (DR5) upregulation through CHOP-dependent and PKC delta-independent mechanism in human malignant tumor cells. Carcinogenesis. 2009; 30:729–736.21. Mérino D, Lalaoui N, Morizot A, Solary E, Micheau O. TRAIL in cancer therapy: present and future challenges. Expert Opin Ther Targets. 2007; 11:1299–1314.22. Fulda S. Tumor-necrosis-factor-related apoptosis-inducing ligand (TRAIL). Adv Exp Med Biol. 2014; 818:167–180.23. Ashkenazi A, Pai RC, Fong S, Leung S, Lawrence DA, Marsters SA, Blackie C, Chang L, McMurtrey AE, Hebert A, DeForge L, Koumenis IL, Lewis D, Harris L, Bussiere J, Koeppen H, Shahrokh Z, Schwall RH. Safety and antitumor activity of recombinant soluble Apo2 ligand. J Clin Invest. 1999; 104:155–162.24. Zhou Y, Tian L, Long L, Quan M, Liu F, Cao J. Casticin potentiates TRAIL-induced apoptosis of gastric cancer cells through endoplasmic reticulum stress. PLoS One. 2013; 8:e58855.25. Kim S, Lee TJ, Leem J, Choi KS, Park JW, Kwon TK. Sanguinarine-induced apoptosis: generation of ROS, down-regulation of Bcl-2, c-FLIP, and synergy with TRAIL. J Cell Biochem. 2008; 104:895–907.26. Yoshida T, Shiraishi T, Nakata S, Horinaka M, Wakada M, Mizutani Y, Miki T, Sakai T. Proteasome inhibitor MG132 induces death receptor 5 through CCAAT/enhancer-binding protein homologous protein. Cancer Res. 2005; 65:5662–5667.27. Shiraishi T, Yoshida T, Nakata S, Horinaka M, Wakada M, Mizutani Y, Miki T, Sakai T. Tunicamycin enhances tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis in human prostate cancer cells. Cancer Res. 2005; 65:6364–6370.28. English AR, Zurek N, Voeltz GK. Peripheral ER structure and function. Curr Opin Cell Biol. 2009; 21:596–602.29. Nishitoh H. CHOP is a multifunctional transcription factor in the ER stress response. J Biochem. 2012; 151:217–219.30. Szewczyk A. The intracellular potassium and chloride channels: properties, pharmacology and function (review). Mol Membr Biol. 1998; 15:49–58.31. Kuum M, Veksler V, Kaasik A. Potassium fluxes across the endoplasmic reticulum and their role in endoplasmic reticulum calcium homeostasis. Cell Calcium. 2015; 58:79–85.32. Liu H, Jiang CC, Lavis CJ, Croft A, Dong L, Tseng HY, Yang F, Tay KH, Hersey P, Zhang XD. 2-Deoxy-D-glucose enhances TRAIL-induced apoptosis in human melanoma cells through XBP-1-mediated up-regulation of TRAIL-R2. Mol Cancer. 2009; 8:122.33. Moon DO, Park SY, Choi YH, Ahn JS, Kim GY. Guggulsterone sensitizes hepatoma cells to TRAIL-induced apoptosis through the induction of CHOP-dependent DR5: involvement of ROS-dependent ER-stress. Biochem Pharmacol. 2011; 82:1641–1650.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Paxilline enhances TRAIL-mediated apoptosis of glioma cells via modulation of c-FLIP, survivin and DR5
- Cytotoxic Effects of Tumor Necrosis Factor-related Apoptosis-inducing Ligand (TRAIL)and its Molecular Mechanism in Human Gastric Cancer Cells
- Vanillin oxime inhibits lung cancer cell proliferation and activates apoptosis through JNK/ERK-CHOP pathway
- Extracellular acidity enhances tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated apoptosis via DR5 in gastric cancer cells
- Troglitazone Increases the Susceptibility to TRAIL-Induced Apoptosis in Thyroid Cancer Cell Lines
