The role of rpoS, hmp, and ssrAB in Salmonella enterica Gallinarum and evaluation of a triple-deletion mutant as a live vaccine candidate in Lohmann layer chickens
- Affiliations
-
- 1College of Veterinary Medicine and Institute of Veterinary Science, Kangwon National University, Chuncheon 200-701, Korea. twhahn@kangwon.ac.kr
- 2Department of Microbiology and Immunology, School of Dentistry, Chosun University, Gwangju 501-759, Korea. isbang@chosun.ac.kr
- KMID: 2164528
- DOI: http://doi.org/10.4142/jvs.2015.16.2.187
Abstract
- Salmonella enterica Gallinarum (SG) causes fowl typhoid (FT), a septicemic disease in avian species. We constructed deletion mutants lacking the stress sigma factor RpoS, the nitric oxide (NO)-detoxifying flavohemoglobin Hmp, and the SsrA/SsrB regulator to confirm the functions of these factors in SG. All gene products were fully functional in wild-type (WT) SG whereas mutants harboring single mutations or a combination of rpoS, hmp, and ssrAB mutations showed hypersusceptibility to H2O2, loss of NO metabolism, and absence of Salmonella pathogenicity island (SPI)-2 expression, respectively. A triple-deletion mutant, SGDelta3 (SGDeltarpoSDeltahmpDeltassrAB), was evaluated for attenuated virulence and protection efficacy in two-week-old Lohmann layer chickens. The SGDelta3 mutant did not cause any mortality after inoculation with either 1 x 10(6) or 1 x 10(8) colony-forming units (CFUs) of bacteria. Significantly lower numbers of salmonellae were recovered from the liver and spleen of chickens inoculated with the SGDelta3 mutant compared to chickens inoculated with WT SG. Vaccination with the SGDelta3 mutant conferred complete protection against challenge with virulent SG on the chickens comparable to the group vaccinated with a conventional vaccine strain, SG9R. Overall, these results indicate that SGDelta3 could be a promising candidate for a live Salmonella vaccine against FT.
Keyword
MeSH Terms
-
Administration, Oral
Animals
Bacterial Proteins/*genetics/immunology
*Chickens
Female
Poultry Diseases/*immunology/microbiology
Salmonella Infections, Animal/*immunology/microbiology
Salmonella Vaccines/administration & dosage/genetics/*immunology
Salmonella enterica/immunology/*physiology
Vaccines, Attenuated/administration & dosage/genetics/immunology
Virulence
Bacterial Proteins
Salmonella Vaccines
Vaccines, Attenuated
Figure
-

Fig. 1 Susceptibility of SG strains to H2O2 or GSNO. Bacterial cells were (A) grown in microtiter plates containing LB broth in the presence (closed symbols) or absence (open symbols) of 250 µM H2O2, and (B) cultured in a microtiter plate containing EG media with (closed symbols) or without (open symbols) 1 mM GSNO. OD600 of the bacteria cultured in the microtiter plates was measured for 24 h at 37℃ with vigorous agitation. Data shown are the mean values ± standard deviation (SD) of three independent cultures. UT: untreated, H2O2: H2O2-treated, GSNO: S-nitrosoglutathione-treated.
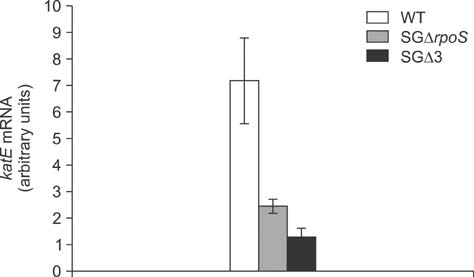
Fig. 2 RpoS-dependent katE transcription in SG. Bacterial cells cultured to log phase in LB media were treated with H2O2 (1 mM) for 30 min. The mRNA levels of katE were measured by quantitative real-time RT-PCR. The values were normalized relative to mRNA levels of a housekeeping gene, gyrB. Data shown are the mean values ± SD of three independent experiments.
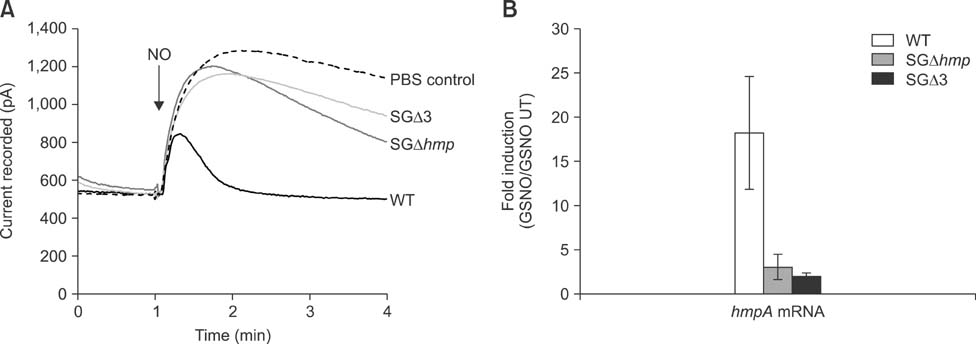
Fig. 3 Role of Hmp in nitric oxide (NO) detoxification of SG. (A) NO consumption of the SG strains was recorded by measuring the NO remaining in PBS containing bacteria after the addition of a fast-releasing NO donor, ProliNONOate (2 µM). A NO-sensitive electrode connected to a free radical analyzer was used to measure NO levels as described in the Materials and Methods section. Data are representative of three independent experiments. (B) WT SG cells were cultured in EG media until OD600 = 0.5. Bacterial transcription was then stopped by adding phenol/ethanol solution to the culture before and 1 h after treatment with GSNO (1 mM). Measured values were normalized relative to mRNA levels of the housekeeping gene gyrB. Data shown are mean fold increases ± SD calculated as the ratio of hmp mRNA levels before and after GSNO treatment for three independent experiments.
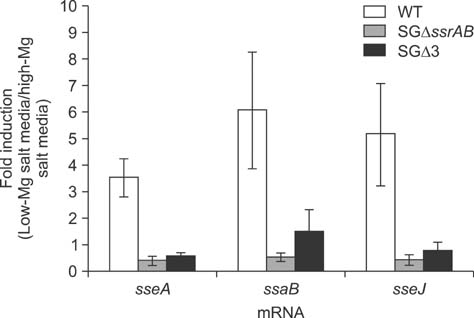
Fig. 4 SPI-2 gene transcription in SG. mRNA levels of sseJ, ssaB, and sseA were measured using total RNA isolated from SG strains cultured under SPI-2-inducing conditions as described in the Materials and Methods section. Measured values were normalized relative to mRNA levels of the housekeeping gene gyrB. Data shown are mean fold increases ± SD calculated as the ratio of mRNA levels before and after the media change for three independent experiments.
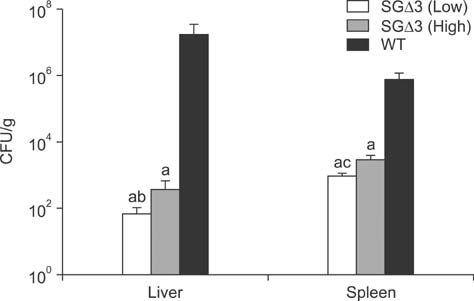
Fig. 5 Attenuation of SGΔ3 in Lohmann layer chickens. The birds were inoculated orally with 1 × 106 (low) or 1 × 108 (high) CFUs of SGΔ3 or 1 × 108 CFUs of WT SG at 2 weeks of age. Bacterial counts in the livers and spleens at 2 weeks post-inoculation are shown as mean values ± SDs of log10 CFU/g for each organ. aIndicates a significant difference (p < 0.0001) between the mutant and WT SG. b,cIndicate a significant difference (p = 0.0008 and p = 0.008) between low and high doses of the SGΔ3 mutant.
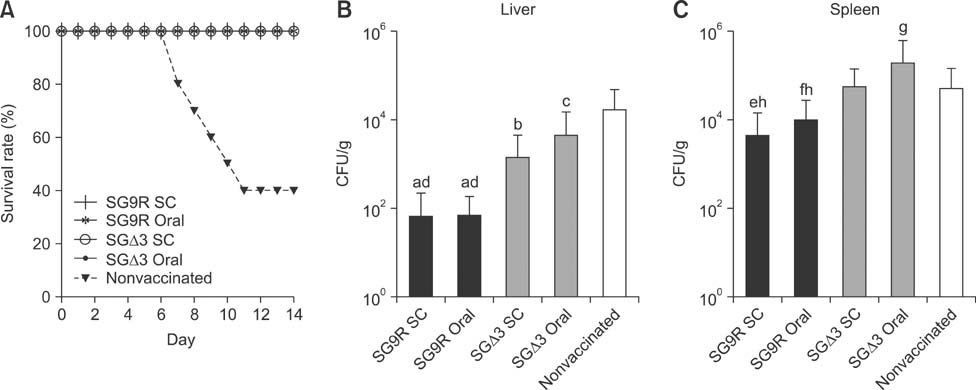
Fig. 6 Protection conferred by SGΔ3 in Lohmann layer chickens. (A) Survival curve for the groups of chickens vaccinated as described in the Materials and Methods section. The chickens were challenged orally at 6 weeks of age with 4 × 108 CFUs of WT SG and observed for 14 days post-challenge (dpc). Bacterial counts in the livers (B) or spleens (C) at 14 dpc are shown as the mean values ± SD of log10 CFU/g for each organ. In the nonvaccinated group, the CFU count was determined for the surviving chickens at 14 dpc. aIndicates a significant difference (p < 0.0001) between SG9R-vaccinated and nonvaccinated chickens. b,cIndicate a significant difference (p = 0.0005 and p = 0.03) between SGΔ3-vaccinated and nonvaccinated chickens. dIndicates a significant difference (p < 0.0001) between SG9R-vaccinated and SGΔ3-vaccinated chickens. e,fIndicate a significant difference (p = 0.0008 and p < 0.01) between SG9R-vaccinated and nonvaccinated chickens. gIndicates a significant difference (p = 0.02) between SGΔ3-vaccinated and nonvaccinated chickens. hIndicates a significant difference (p < 0.001) between SG9R-vaccinated and SGΔ3-vaccinated chickens.
Reference
-
1. Bang IS, Frye JG, McClelland M, Velayudhan J, Fang FC. Alternative sigma factor interactions in Salmonella: σE and σH promote antioxidant defenses by enhancing σS levels. Mol Microbiol. 2005; 56:811–823.
Article2. Bang IS, Liu L, Vazquez-Torres A, Crouch ML, Stamler JS, Fang FC. Maintenance of nitric oxide and redox homeostasis by the Salmonella flavohemoglobin Hmp. J Biol Chem. 2006; 281:28039–28047.
Article3. Barrow PA, Freitas Neto OC. Pullorum disease and fowl typhoid-new thoughts on old diseases: a review. Avian Pathol. 2011; 40:1–13.
Article4. Bouzoubaa K, Nagaraja KV, Kabbaj FZ, Newman JA, Pomeroy BS. Feasibility of using proteins from Salmonella gallinarum vs. 9R live vaccine for the prevention of fowl typhoid in chickens. Avian Dis. 1989; 33:385–391.
Article5. Boyen F, Pasmans F, Van Immerseel F, Morgan E, Botteldoorn N, Heyndrickx M, Volf J, Favoreel H, Hernalsteens JP, Ducatelle R, Haesebrouck F. A limited role for SsrA/B in persistent Salmonella Typhimurium infections in pigs. Vet Microbiol. 2008; 128:364–373.
Article6. Coynault C, Robbe-Saule V, Norel F. Virulence and vaccine potential of Salmonella typhimurium mutants deficient in the expression of the RpoS (σS) regulon. Mol Microbiol. 1996; 22:149–160.
Article7. Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A. 2000; 97:6640–6645.
Article8. Deiwick J, Nikolaus T, Erdogan S, Hensel M. Environmental regulation of Salmonella pathogenicity island 2 gene expression. Mol Microbiol. 1999; 31:1759–1773.
Article9. Fang FC, Libby SJ, Buchmeier NA, Loewen PC, Switala J, Harwood J, Guiney DG. The alternative σ factor KatF (RpoS) regulates Salmonella virulence. Proc Natl Acad Sci U S A. 1992; 89:11978–11982.10. Foley SL, Johnson TJ, Ricke SC, Nayak R, Danzeisen J. Salmonella pathogenicity and host adaptation in chicken-associated serovars. Microbiol Mol Biol Rev. 2013; 77:582–607.
Article11. Forrester MT, Foster MW. Protection from nitrosative stress: a central role for microbial flavohemoglobin. Free Radic Biol Med. 2012; 52:1620–1633.
Article12. Hart TW. Some observations concerning the S-nitroso and S-phenylsulphonyl derivatives of L-cysteine and glutathione. Tetrahedron Lett. 1985; 26:2013–2016.
Article13. Jeong JH, Song M, Park SI, Cho KO, Rhee JH, Choy HE. Salmonella enterica serovar Gallinarum requires ppGpp for internalization and survival in animal cells. J Bacteriol. 2008; 190:6340–6350.
Article14. Jones MA, Wigley P, Page KL, Hulme SD, Barrow PA. Salmonella enterica serovar Gallinarum requires the Salmonella pathogenicity island 2 type III secretion system but not the Salmonella pathogenicity island 1 type III secretion system for virulence in chickens. Infect Immun. 2001; 69:5471–5476.
Article15. Kazmierczak MJ, Wiedmann M, Boor KJ. Alternative sigma factors and their roles in bacterial virulence. Microbiol Mol Biol Rev. 2005; 69:527–543.
Article16. Kwon HJ, Cho SH. Pathogenicity of SG 9R, a rough vaccine strain against fowl typhoid. Vaccine. 2011; 29:1311–1318.
Article17. Löber S, Jäckel D, Kaiser N, Hensel M. Regulation of Salmonella pathogenicity island 2 genes by independent environmental signals. Int J Med Microbiol. 2006; 296:435–447.
Article18. Morgan E, Campbell JD, Rowe SC, Bispham J, Stevens MP, Bowen AJ, Barrow PA, Maskell DJ, Wallis TS. Identification of host-specific colonization factors of Salmonella enterica serovar Typhimurium. Mol Microbiol. 2004; 54:994–1010.
Article19. Pasmans F, Van Immerseel F, Heyndrickx M, Martel A, Godard C, Wildemauwe C, Ducatelle R, Haesebrouck F. Host adaptation of pigeon isolates of Salmonella enterica subsp. enterica serovar Typhimurium variant Copenhagen phage type 99 is associated with enhanced macrophage cytotoxicity. Infect Immun. 2003; 71:6068–6074.
Article20. Robbe-Saule V, Algorta G, Rouilhac I, Norel F. Characterization of the RpoS status of clinical isolates of Salmonella enterica. Appl Environ Microbiol. 2003; 69:4352–4358.
Article21. Seo YS, Lee SH, Shin EK, Kim SJ, Jung R, Hahn TW. Pulsed-field gel electrophoresis genotyping of Salmonella gallinarum and comparison with random amplified polymorphic DNA. Vet Microbiol. 2006; 115:349–357.
Article22. Silva EN, Snoeyenbos GH, Weinack OM, Smyser CF. Studies on the use of 9R strain of Salmonella gallinarum as a vaccine in chickens. Avian Dis. 1981; 25:38–52.
Article23. Smith HW. The immunity to Salmonella gallinarum infection in chickens produced by live cultures of members of the Salmonella genus. J Hyg (Lond). 1956; 54:433–439.
Article24. Van Immerseel F, Studholme DJ, Eeckhaut V, Heyndrickx M, Dewulf J, Dewaele I, Van Hoorebeke S, Haesebrouck F, Van Meirhaeghe H, Ducatelle R, Paszkiewicz K, Titball RW. Salmonella Gallinarum field isolates from laying hens are related to the vaccine strain SG9R. Vaccine. 2013; 31:4940–4945.
Article25. Waterman SR, Holden DW. Functions and effectors of the Salmonella pathogenicity island 2 type III secretion system. Cell Microbiol. 2003; 5:501–511.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Protection Against Salmonella Typhimurium, Salmonella Gallinarum, and Salmonella Enteritidis Infection in Layer Chickens Conferred by a Live Attenuated Salmonella Typhimurium Strain
- Protective efficacy and immune responses by homologous prime-booster immunizations of a novel inactivated Salmonella Gallinarum vaccine candidate
- Comparative evaluation of the murine immune responses to Salmonella enterica serovars Enteritidis, Gallinarum and Typhimurium infection
- Development and evaluation of protective capacity of Salmonella Enteritidis polyphosphate kinase-deleted and temperature-sensitive mutant
- Establishment of a live vaccine strain against fowl typhoid and paratyphoid
