Gap Junction Contributions to the Goldfish Electroretinogram at the Photopic Illumination Level
- Affiliations
-
- 1Natural Sciences Section, Department of Medical Lifescience, College of Medicine, The Catholic University of Korea, Seoul 137-701, Korea. cjung@catholic.ac.kr
- KMID: 1768017
- DOI: http://doi.org/10.4196/kjpp.2012.16.3.219
Abstract
- Understanding how the b-wave of the electroretinogram (ERG) is generated by full-field light stimulation is still a challenge in visual neuroscience. To understand more about the origin of the b-wave, we studied the contributions of gap junctions to the ERG b-wave. Many types of retinal neurons are connected to similar and different neighboring neurons through gap junctions. The photopic (cone-dominated) ERG, stimulated by a small light beam, was recorded from goldfish (Carassius auratus) using a corneal electrode. Data were obtained before and after intravitreal injection of agents into the eye under a photopic illumination level. Several agents were used to affect gap junctions, such as dopamine D1 and D2 receptor agonists and antagonists, a nitric oxide (NO) donor, a nitric oxide synthase (NOS) inhibitor, the gap junction blocker meclofenamic acid (MFA), and mixtures of these agents. The ERG b-waves, which were enhanced by MFA, sodium nitroprusside (SNP), SKF 38393, and sulpiride, remained following application of a further injection of a mixture with MFA. The ERG b-waves decreased following NG-nitro-L-arginine methyl ester (L-NAME), SCH 23390, and quinpirole administration but were enhanced by further injection of a mixture with MFA. These results indicate that gap junction activity influences b-waves of the ERG related to NO and dopamine actions.
Keyword
MeSH Terms
-
2,3,4,5-Tetrahydro-7,8-dihydroxy-1-phenyl-1H-3-benzazepine
Benzazepines
Dopamine
Electrodes
Eye
Gap Junctions
Goldfish
Humans
Intravitreal Injections
Light
Lighting
Meclofenamic Acid
Neurons
Neurosciences
NG-Nitroarginine Methyl Ester
Nitric Oxide
Nitric Oxide Synthase
Nitroprusside
Quinpirole
Retinal Neurons
Sulpiride
Tissue Donors
2,3,4,5-Tetrahydro-7,8-dihydroxy-1-phenyl-1H-3-benzazepine
Benzazepines
Dopamine
Meclofenamic Acid
NG-Nitroarginine Methyl Ester
Nitric Oxide
Nitric Oxide Synthase
Nitroprusside
Quinpirole
Sulpiride
Figure
-
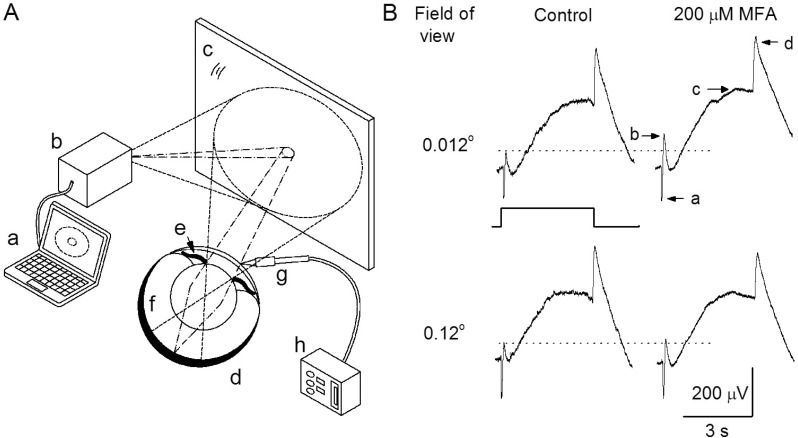
Fig. 1 A schematic drawing, as seen from the side, of the overall process of the light stimulus for electroretinogram (ERG) measurements and consequent responses. (A) The dashed and dotted lines show a sketch of the ray paths of the smaller and larger white circle beams emitted from the projector, respectively. a, computer; b, beam projector; c, surface mirror; d, eye ball; e, iris; f, retina (shaded area); g, recording electrode; h, differential amplifier. (B) The ERG responses were evoked by small and large sizes of circle light beam before (upper and down, left) and after (upper and down, right) intravitreal injection of 200 µM MFA. The small and large sizes of the light beam imaged on the surface of the mirror were 6 mm and 60 mm in diameter corresponding to the values of 0.012° and 0.12° in the field of view. Light stimulus was a 4-s presented by the square wave. The components of ERG a-, b-, c-, and d-waves are denoted as a, b, c, and d.
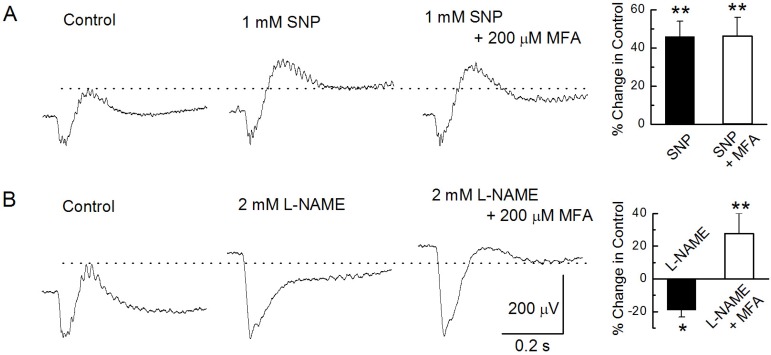
Fig. 2 Modulation of the b-wave. (A) The b-wave was recorded under the control condition, after injecting 1 mM SNP, and after injecting a mixture of 1 mM SNP and 200 µM MFA. (B) The b-wave was recorded under the control condition, after injecting 2 mM L-NAME, and after injecting a mixture of 2 mM L-NAME and 200 µM MFA. In bar graphs end of the each low line, the mean peak amplitudes of the b-wave by drugs are plotted as a percentage of the peak response under control condition. In this and all subsequent figures, each bar represents the mean±standard error and single (p<0.05) and double stars (p<0.01) above the data column indicate significant and very significant statistical differences, respectively. See text for details of the statistical comparisons.
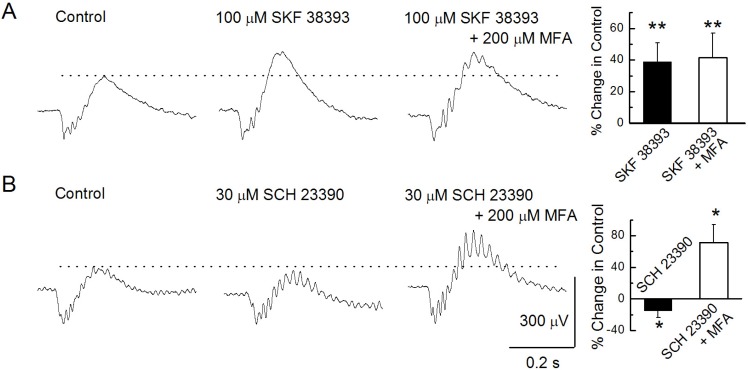
Fig. 3 Modulation of b-waves. (A) b-waves were recorded under the control condition, after injecting 100 µM SKF 38393, and after injecting a mixture of 100 µM SKF 38393 and 200 µM MFA. (B) b-waves were recorded under the control condition, after injecting 30 µM SCH 23390, and after injecting a mixture of 30 µM SCH 23390 and 200 µM MFA.
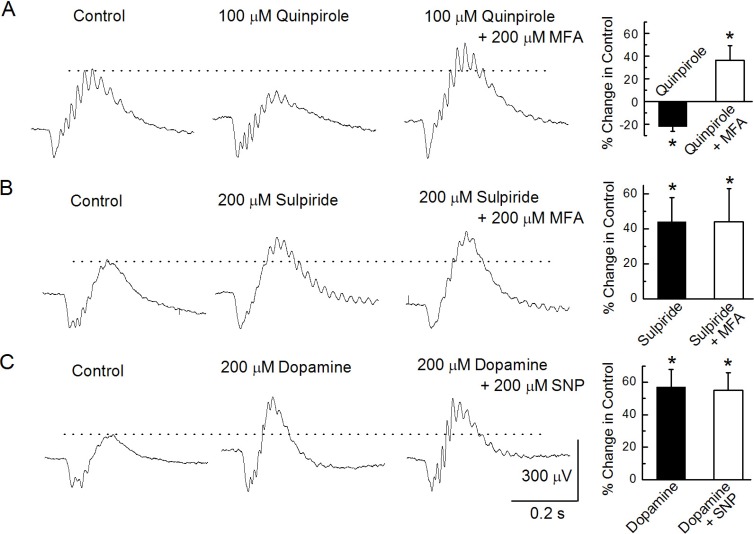
Fig. 4 Modulation of b-waves. (A) b-waves were recorded under the control condition, after injecting 100 µM quinpirole, and after injecting a mixture of 100 µM quinpirole and 200 µM MFA. (B) b-waves were recorded under the control condition, after injecting 200 µM sulpiride, and after injecting a mixture of 200 µM sulpiride and 200 µM MFA. (C) b-waves were recorded under the control condition, after injecting 200 µM dopamine, and after injecting a mixture of 200 µM dopamine and 200 µM MFA.
Cited by 1 articles
-
Bisphenol A and 4-
tert -Octylphenol Inhibit Cx46 Hemichannel Currents
Seunghoon Oh
Korean J Physiol Pharmacol. 2015;19(1):73-79. doi: 10.4196/kjpp.2015.19.1.73.
Reference
-
1. Bloomfield SA, Völgyi B. The diverse functional roles and regulation of neuronal gap junctions in the retina. Nat Rev Neurosci. 2009; 10:495–506. PMID: 19491906.
Article2. Werblin FS, Dowling JE. Organization of the retina of the mudpuppy, Necturus maculosus. II. Intracellular recording. J Neurophysiol. 1969; 32:339–355. PMID: 4306897.
Article3. Kaneko A. Receptive field organization of bipolar and amacrine cells in the goldfish retina. J Physiol. 1973; 235:133–153. PMID: 4778132.
Article4. Naka KI, Nye PW. Role of horizontal cells in organization of the catfish retinal receptive field. J Neurophysiol. 1971; 34:785–801. PMID: 5097157.
Article5. Marchiafava PL. Horizontal cells influence membrane potential of bipolar cells in the retina of the turtle. Nature. 1978; 275:141–142. PMID: 692683.
Article6. Arai I, Tanaka M, Tachibana M. Active roles of electrically coupled bipolar cell network in the adult retina. J Neurosci. 2010; 30:9260–9270. PMID: 20610761.
Article7. Cook JE, Becker DL. Gap junctions in the vertebrate retina. Microsc Res Tech. 1995; 31:408–419. PMID: 8534902.
Article8. Marc RE, Liu WL, Muller JF. Gap junctions in the inner plexiform layer of the goldfish retina. Vision Res. 1988; 28:9–24. PMID: 3414003.
Article9. Saito T, Kujiraoka T. Characteristics of bipolar-bipolar coupling in the carp retina. J Gen Physiol. 1988; 91:275–287. PMID: 3373179.10. DeVries SH, Schwartz EA. Modulation of an electrical synapse between solitary pairs of catfish horizontal cells by dopamine and second messengers. J Physiol. 1989; 414:351–375. PMID: 2558170.
Article11. DeVries SH, Schwartz EA. Hemi-gap-junction channels in solitary horizontal cells of the catfish retina. J Physiol. 1992; 445:201–230. PMID: 1380084.
Article12. Lu C, McMahon DG. Modulation of hybrid bass retinal gap junctional channel gating by nitric oxide. J Physiol. 1997; 499:689–699. PMID: 9130165.
Article13. Lasater EM, Dowling JE. Dopamine decreases conductance of the electrical junctions between cultured retinal horizontal cells. Proc Natl Acad Sci USA. 1985; 82:3025–3029. PMID: 3857632.
Article14. Piccolino M, Neyton J, Gerschenfeld HM. Decrease of gap junction permeability induced by dopamine and cyclic adenosine 3':5'-monophosphate in horizontal cells of turtle retina. J Neurosci. 1984; 4:2477–2488. PMID: 6092564.
Article15. Hedden WL Jr, Dowling JE. The interplexiform cell system. II. Effects of dopamine on goldfish retinal neurones. Proc R Soc Lond B Biol Sci. 1978; 201:27–55. PMID: 27790.16. Murakami M, Miyachi EI, Takahadhi KI. Modulation of gap juctions between horizontal cells by second messengers. Prog Retinal Eye Res. 1995; 14:197–221.17. McMahon DG, Brown DR. Modulation of gap-junction channel gating at zebrafish retinal electrical synapses. J Neurophysiol. 1994; 72:2257–2268. PMID: 7533830.
Article18. Baldridge WH, Ball AK, Miller RG. Dopaminergic regulation of horizontal cell gap junction particle density in goldfish retina. J Comp Neurol. 1987; 265:428–436. PMID: 3693614.
Article19. Stell WK. Inputs to bipolar cell dendrites in goldfish retina. Sens Processes. 1978; 2:339–349. PMID: 755290.20. Saito T, Kondo H, Toyoda J. Rod and cone signals in the on-center bipolar cell: their different ionic mechanisms. Vision Res. 1978; 18:591–595. PMID: 664343.
Article21. Malchow RP, Yazulla S. Separation and light adaptation of rod and cone signals in the retina of the goldfish. Vision Res. 1986; 26:1655–1666. PMID: 3617507.
Article22. Hughes A, Saszik S, Bilotta J, Demarco PJ Jr, Patterson WF 2nd. Cone contributions to the photopic spectral sensitivity of the zebrafish ERG. Vis Neurosci. 1998; 15:1029–1037. PMID: 9839967.
Article23. Hood DC, Finkelstein MA. Boff KR, Kaufman L, Thomas JP, editors. Sensitivity to light. Handbook of Perception and Human Performance. 1986. New York: Wiley;p. 5-1–5-66.24. Kim SH, Jung CS. The role of the pattern edge in goldfish visual motion detection. Korean J Physiol Pharmacol. 2010; 14:413–417. PMID: 21311683.
Article25. Liu CR, Xu L, Zhong YM, Li RX, Yang XL. Expression of connexin 35/36 in retinal horizontal and bipolar cells of carp. Neuroscience. 2009; 164:1161–1169. PMID: 19778581.
Article26. O'Brien J, al-Ubaidi MR, Ripps H. Connexin 35: a gap-junctional protein expressed preferentially in the skate retina. Mol Biol Cell. 1996; 7:233–243. PMID: 8688555.27. Condorelli DF, Parenti R, Spinella F, Trovato Salinaro A, Belluardo N, Cardile V, Cicirata F. Cloning of a new gap junction gene (Cx36) highly expressed in mammalian brain neurons. Eur J Neurosci. 1998; 10:1202–1208. PMID: 9753189.
Article28. Veruki ML, Hartveit E. Meclofenamic acid blocks electrical synapses of retinal AII amacrine and on-cone bipolar cells. J Neurophysiol. 2009; 101:2339–2347. PMID: 19279153.
Article29. Pan F, Mills SL, Massey SC. Screening of gap junction antagonists on dye coupling in the rabbit retina. Vis Neurosci. 2007; 24:609–618. PMID: 17711600.
Article30. Tomita T, Yanagida T. Origins of the ERG waves. Vision Res. 1981; 21:1703–1707. PMID: 7336605.
Article31. Koriyama Y, Yasuda R, Homma K, Mawatari K, Nagashima M, Sugitani K, Matsukawa T, Kato S. Nitric oxide-cGMP signaling regulates axonal elongation during optic nerve regeneration in the goldfish in vitro and in vivo. J Neurochem. 2009; 110:890–901. PMID: 19457064.32. Villani L, Guarnieri T. Localization of nitric oxide synthase in the goldfish retina. Brain Res. 1996; 743:353–356. PMID: 9017268.
Article33. Liepe BA, Stone C, Koistinaho J, Copenhagen DR. Nitric oxide synthase in Müller cells and neurons of salamander and fish retina. J Neurosci. 1994; 14:7641–7654. PMID: 7527846.34. Yazulla S, Lin ZS. Differential effects of dopamine depletion on the distribution of [3H]SCH 23390 and [3H]spiperone binding sites in the goldfish retina. Vision Res. 1995; 35:2409–2414. PMID: 8594810.35. Harsanyi K, Mangel SC. Activation of a D2 receptor increases electrical coupling between retinal horizontal cells by inhibiting dopamine release. Proc Natl Acad Sci USA. 1992; 89:9220–9224. PMID: 1357661.
Article36. Van Buskirk R, Dowling JE. Isolated horizontal cells from carp retina demonstrate dopamine-dependent accumulation of cyclic AMP. Proc Natl Acad Sci USA. 1981; 78:7825–7829. PMID: 6278491.
Article37. Lasater EM. Retinal horizontal cell gap junctional conductance is modulated by dopamine through a cyclic AMP-dependent protein kinase. Proc Natl Acad Sci USA. 1987; 84:7319–7323. PMID: 2823257.
Article38. Huppé-Gourgues F, Coudé G, Lachapelle P, Casanova C. Effects of the intravitreal administration of dopaminergic ligands on the b-wave amplitude of the rabbit electroretinogram. Vision Res. 2005; 45:137–145. PMID: 15581915.
Article39. Hedden WL Jr, Dowling JE. The interplexiform cell system. II. Effects of dopamine on goldfish retinal neurones. Proc R Soc Lond B Biol Sci. 1978; 201:27–55. PMID: 27790.40. Dowling JE, Ehinger B. Synaptic organization of the dopaminergic neurons in the rabbit retina. J Comp Neurol. 1978; 180:203–220. PMID: 207745.
Article41. Dowling JE, Ehinger B. The interplexiform cell system. I. Synapses of the dopaminergic neurons of the goldfish retina. Proc R Soc Lond B Biol Sci. 1978; 201:7–26. PMID: 27792.42. Hampton CK, Redburn DA. Autoradiographic analysis of 3H-glutamate, 3H-dopamine, and 3H-GABA accumulation in rabbit retina after kainic acid treatment. J Neurosci Res. 1983; 9:239–251. PMID: 6133958.43. Yazulla S, Zucker CL. Synaptic organization of dopaminergic interplexiform cells in the goldfish retina. Vis Neurosci. 1988; 1:13–29. PMID: 2908724.
Article44. Djamgoz MB, Wagner HJ. Localization and function of dopamine in the adult vertebrate retina. Neurochem Int. 1992; 20:139–191. PMID: 1304857.
Article45. Negishi K, Teranishi T, Kato S. The dopamine system of the teleost fish retina. Prog Ret Res. 1990; 9:1–48.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Role of the Pattern Edge in Goldfish Visual Motion Detection
- Photopic Negative Response (PhNR) in Normal Subjects
- Photopic Electroretinogram in Adult Diabetics
- The Change of Contrast Sensitivity in Amblyopic Patient after Occlusion Therapy using ACV
- ON and OFF Responses of the Electroretinogram in Patients with Glaucoma
