Altered Metabolic Phenotypes and Hypothalamic Neuronal Activity Triggered by Sodium-Glucose Cotransporter 2 Inhibition
- Affiliations
-
- 1Division of Life Sciences, College of Life Sciences and Bioengineering, Incheon National University, Incheon, Korea
- 2Department of Biological Science, University of Ulsan, Ulsan, Korea
- 3Division of Sport Science, College of Arts & Physical Education, Incheon National University, Incheon, Korea
- 4Research Center of Brain-Machine Interface, Incheon National University, Incheon, Korea
- 5Department of Nano-Bioengineering, Incheon National University, Incheon, Korea
- 6Department of Internal Medicine, Ulsan University Hospital, University of Ulsan College of Medicine, Ulsan, Korea
- KMID: 2548155
- DOI: http://doi.org/10.4093/dmj.2022.0261
Abstract
- Background
Sodium-glucose cotransporter 2 (SGLT-2) inhibitors are currently used to treat patients with diabetes. Previous studies have demonstrated that treatment with SGLT-2 inhibitors is accompanied by altered metabolic phenotypes. However, it has not been investigated whether the hypothalamic circuit participates in the development of the compensatory metabolic phenotypes triggered by the treatment with SGLT-2 inhibitors.
Methods
Mice were fed a standard diet or high-fat diet and treated with dapagliflozin, an SGLT-2 inhibitor. Food intake and energy expenditure were observed using indirect calorimetry system. The activity of hypothalamic neurons in response to dapagliflozin treatment was evaluated by immunohistochemistry with c-Fos antibody. Quantitative real-time polymerase chain reaction was performed to determine gene expression patterns in the hypothalamus of dapagliflozin-treated mice.
Results
Dapagliflozin-treated mice displayed enhanced food intake and reduced energy expenditure. Altered neuronal activities were observed in multiple hypothalamic nuclei in association with appetite regulation. Additionally, we found elevated immunosignals of agouti-related peptide neurons in the paraventricular nucleus of the hypothalamus.
Conclusion
This study suggests the functional involvement of the hypothalamus in the development of the compensatory metabolic phenotypes induced by SGLT-2 inhibitor treatment.
Keyword
Figure
-

Fig. 1. (A) Experiment 1: To examine the effects of dapagliflozin (sodium-glucose cotransporter 2 inhibitor, Sigma-Aldrich) on metabolic phenotypes, mice were randomly divided into two groups and fed a standard diet (SD, 10% calories from fat, Research Diets Inc.). For treatment, SD-fed mice were administered with vehicle (0.9% saline, oral injection) or dapagliflozin (1 mg/kg/day, oral injection) for 5 weeks. (B) Experiment 2: To test the effects of dapagliflozin on metabolic phenotypes during overnutrition, mice were fed a high-fat diet (HFD, 60% of calories from fat, Research Diets Inc.) for 13 weeks. Eight weeks after the HFD treatment, mice were treated with dapagliflozin (1 mg/kg/day, oral injection), while HFD treatment was continued.

Fig. 2. Effect of dapagliflozin on variations in food intake and body weight. Body weight and food intake were evaluated in vehicle- or dapagliflozin-treated mice under standard diet (SD) or high-fat diet (HFD) feeding conditions using an indirect calorimetry system. No significant alterations in body weight were found in dapagliflozin-treated mice under (A) SD or (B) HFD feeding conditions. (C) Cumulative food intake over 48 hours was enhanced in dapagliflozin-treated mice under SD feeding conditions. (D) Meal frequency during the dark period and (E) meal size during the light period were higher in dapagliflozin-treated mice than those in vehicle-treated mice under SD feeding conditions. (F) Cumulative food intake was increased in dapagliflozin-treated mice compared with that in vehicle-treated mice under HFD feeding conditions. (G) No significant difference in meal frequency and (H) increased meal size were observed in dapagliflozin-treated mice under HFD feeding conditions. Data are presented as the mean±standard error of the mean (n=6 mice for SD group, n=7 mice for HFD group). aP<0.05.

Fig. 3. Treatment with dapagliflozin leads to a decrease in energy expenditure in high-fat diet (HFD)-fed mice. The patterns of energy expenditure were measured in vehicle- or dapagliflozin-treated mice using an indirect calorimetry system. Under standard diet (SD) feeding conditions, dapagliflozin-treated mice displayed no alteration in (A) oxygen consumption (VO2), (B) carbon dioxide emission (VCO2), (C) respiratory exchange ratio (RER), or (D) energy expenditure compared with control mice. Under HFD feeding conditions, dapagliflozin-treated mice showed reduced (E) VO2, (F) VCO2, and (H) energy expenditure with no alteration in (G) RER compared with control mice. Data are presented as the mean±standard error of the mean (n=6 mice for SD group, n=8 mice for HFD group). aP<0.05, bP<0.01.

Fig. 4. Treatment with dapagliflozin alters the neuronal activities in the hypothalamic nuclei of mice. (A) Representative images showing immunosignals of c-Fos-positive cells in hypothalamic nuclei of vehicle- or dapagliflozin-treated mice under standard diet (SD) feeding conditions. (B) Dapagliflozin treatment resulted in an increasing number of c-Fos-positive cells in the hypothalamic arcuate nucleus (Arc) and lateral hypothalamus (Lh) and a decreasing number of c-Fos-positive cells in the hypothalamic ventromedial hypothalamus (Vmh) and paraventricular nucleus (Pvn). (C) Representative images showing immunosignals of c-Fos-positive cells in the hypothalamic nuclei of vehicle- or dapagliflozin-treated mice under high-fat diet (HFD) feeding conditions. (D) Dapagliflozin treatment resulted in an increase in c-Fos-positive cells in the hypothalamic Arc and a decrease in c-Fos-positive cells in the hypothalamic Lh in HFD-fed mice. Data are presented as the mean±standard error of the mean (n=5 mice for SD-vehicle-treated group, n=4 mice for SD-dapagliflozin-treated group, n=6 mice for HFD-vehicle-treated group; HFD-dapagliflozin-treated group). Scale bar=100 µm. Dmh, dorsomedial hypothalamus. aP<0.05, bP<0.01.
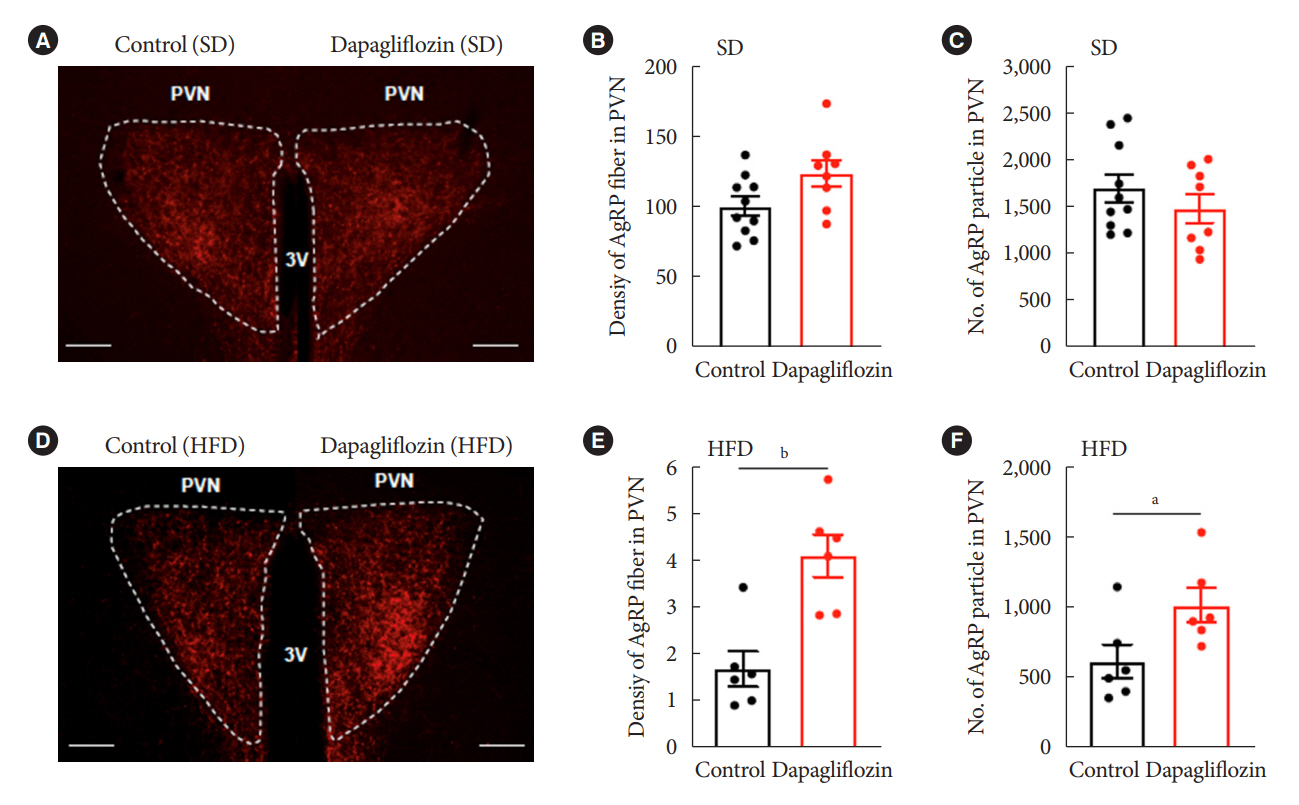
Fig. 5. Dapagliflozin treatment enhances immunosignals of agouti-related peptide (Agrp) in the paraventricular nucleus (Pvn) of high-fat diet (HFD)-fed mice. (A) Representative images showing immunosignals of Agrp fibers in the hypothalamic Pvn of standard diet (SD)-fed mice. Treatment with dapagliflozin did not alter (B) fiber density and (C) particle numbers of Agrp-positive immunosignals in the Pvn of SD-fed mice. (D) Representative image showing immunosignals of Agrp fibers in the hypothalamic Pvn of HFD-fed mice. An increase in the (E) fiber density and (F) particle numbers of Agrp-positive immunosignals was observed in the Pvn of dapagliflozin-treated mice under HFD feeding conditions. Data are presented as the mean±standard error of the mean (n=5 mice for SD-vehicle-treated group, n=4 mice for SD-dapagliflozin-treated group, n=6 mice for HFD-vehicle-treated group; HFD-dapagliflozin-treated group). Scale bar=100 µm. aP<0.05, bP<0.01.

Fig. 6. The expression patterns of hypothalamic mRNA in response to dapagliflozin treatment. The levels of mRNA involved in appetite regulation were analyzed in the hypothalamus of vehicle- or dapagliflozin-treated mice under standard diet (SD) or high-fat diet (HFD) feeding conditions. (A) The agouti-related peptide (Agrp) mRNA level shows an elevation in the hypothalamus of dapagliflozin-treated mice compared with that in control mice under SD feeding conditions. The mRNA levels of (B) pro-opiomelanocortin (Pomc) and (C) neuropeptide Y (Npy) did not respond to dapagliflozin treatment under SD or HFD feeding conditions. Dapagliflozin treatment did not affect the mRNA expression of (D) suppressor of cytokine signaling 3 (Socs3) and (E) forkhead box protein O1 (Foxo1) in the hypothalamus of SD- or HFD-fed mice. Data are presented as the mean±standard error of the mean (n=4 mice for SD-vehicle-treated group; HFD-vehicle-treated group; n=5 mice for SD-dapagliflozin-treated group; HFD-dapagliflozin-treated group). aP<0.05.
Reference
-
1. Abdul-Ghani MA, Norton L, DeFronzo RA. Renal sodium-glucose cotransporter inhibition in the management of type 2 diabetes mellitus. Am J Physiol Renal Physiol. 2015; 309:F889–900.
Article2. Wright EM, Loo DD, Hirayama BA. Biology of human sodium glucose transporters. Physiol Rev. 2011; 91:733–94.
Article3. Rieg T, Masuda T, Gerasimova M, Mayoux E, Platt K, Powell DR, et al. Increase in SGLT1-mediated transport explains renal glucose reabsorption during genetic and pharmacological SGLT2 inhibition in euglycemia. Am J Physiol Renal Physiol. 2014; 306:F188–93.
Article4. Neuwirt H, Burtscher A, Cherney D, Mayer G, Ebenbichler C. Tubuloglomerular feedback in renal glucosuria: mimicking long-term SGLT-2 inhibitor therapy. Kidney Med. 2019; 2:76–9.
Article5. Vallon V, Thomson SC. Targeting renal glucose reabsorption to treat hyperglycaemia: the pleiotropic effects of SGLT2 inhibition. Diabetologia. 2017; 60:215–25.
Article6. Zaccardi F, Webb DR, Htike ZZ, Youssef D, Khunti K, Davies MJ. Efficacy and safety of sodium-glucose co-transporter-2 inhibitors in type 2 diabetes mellitus: systematic review and network meta-analysis. Diabetes Obes Metab. 2016; 18:783–94.
Article7. Thomas MC, Cherney DZI. The actions of SGLT2 inhibitors on metabolism, renal function and blood pressure. Diabetologia. 2018; 61:2098–107.
Article8. Komiya C, Tsuchiya K, Shiba K, Miyachi Y, Furuke S, Shimazu N, et al. Ipragliflozin improves hepatic steatosis in obese mice and liver dysfunction in type 2 diabetic patients irrespective of body weight reduction. PLoS One. 2016; 11:e0151511.
Article9. Saper CB, Lowell BB. The hypothalamus. Curr Biol. 2014; 24:R1111–6.
Article10. Dietrich MO, Horvath TL. Hypothalamic control of energy balance: insights into the role of synaptic plasticity. Trends Neurosci. 2013; 36:65–73.
Article11. Lopez-Gambero AJ, Martinez F, Salazar K, Cifuentes M, Nualart F. Brain glucose-sensing mechanism and energy homeostasis. Mol Neurobiol. 2019; 56:769–96.
Article12. Horvath TL, Sarman B, Garcia-Caceres C, Enriori PJ, Sotonyi P, Shanabrough M, et al. Synaptic input organization of the melanocortin system predicts diet-induced hypothalamic reactive gliosis and obesity. Proc Natl Acad Sci U S A. 2010; 107:14875–80.
Article13. Cansell C, Denis RG, Joly-Amado A, Castel J, Luquet S. Arcuate AgRP neurons and the regulation of energy balance. Front Endocrinol (Lausanne). 2012; 3:169.
Article14. Kusakabe T, Yokota S, Shimizu M, Inoue T, Tanaka M, Ohue-Kitano R, et al. Differential effects of sodium-glucose cotransporter 2 inhibitor and low-carbohydrate diet on body composition and metabolic profile in obese diabetic db/db mice. BMJ Open Diabetes Res Care. 2020; 8:e001303.15. Inoue H, Morino K, Ugi S, Tanaka-Mizuno S, Fuse K, Miyazawa I, et al. Ipragliflozin, a sodium-glucose cotransporter 2 inhibitor, reduces bodyweight and fat mass, but not muscle mass, in Japanese type 2 diabetes patients treated with insulin: a randomized clinical trial. J Diabetes Investig. 2019; 10:1012–21.
Article16. Ferrannini G, Hach T, Crowe S, Sanghvi A, Hall KD, Ferrannini E. Energy balance after sodium-glucose cotransporter 2 inhibition. Diabetes Care. 2015; 38:1730–5.
Article17. Chiba Y, Yamada T, Tsukita S, Takahashi K, Munakata Y, Shirai Y, et al. Dapagliflozin, a sodium-glucose co-transporter 2 inhibitor, acutely reduces energy expenditure in BAT via neural signals in mice. PLoS One. 2016; 11:e0150756.
Article18. van Ruiten CC, Veltman DJ, Schrantee A, van Bloemendaal L, Barkhof F, Kramer MH, et al. Effects of dapagliflozin and combination therapy with exenatide on food-cue induced brain activation in patients with type 2 diabetes. J Clin Endocrinol Metab. 2022; 107:e2590–9.
Article19. Schwartz MW, Woods SC, Porte D Jr, Seeley RJ, Baskin DG. Central nervous system control of food intake. Nature. 2000; 404:661–71.
Article20. Nguyen T, Wen S, Gong M, Yuan X, Xu D, Wang C, et al. Dapagliflozin activates neurons in the central nervous system and regulates cardiovascular activity by inhibiting SGLT-2 in mice. Diabetes Metab Syndr Obes. 2020; 13:2781–99.21. Takeda K, Ono H, Ishikawa K, Ohno T, Kumagai J, Ochiai H, et al. Central administration of sodium-glucose cotransporter-2 inhibitors increases food intake involving adenosine monophosphate-activated protein kinase phosphorylation in the lateral hypothalamus in healthy rats. BMJ Open Diabetes Res Care. 2021; 9:e002104.
Article22. Kim JD, Leyva S, Diano S. Hormonal regulation of the hypothalamic melanocortin system. Front Physiol. 2014; 5:480.
Article23. Valassi E, Scacchi M, Cavagnini F. Neuroendocrine control of food intake. Nutr Metab Cardiovasc Dis. 2008; 18:158–68.
Article24. Sobrino Crespo C, Perianes Cachero A, Puebla Jimenez L, Barrios V, Arilla Ferreiro E. Peptides and food intake. Front Endocrinol (Lausanne). 2014; 5:58.
Article25. Enriori PJ, Evans AE, Sinnayah P, Jobst EE, Tonelli-Lemos L, Billes SK, et al. Diet-induced obesity causes severe but reversible leptin resistance in arcuate melanocortin neurons. Cell Metab. 2007; 5:181–94.
Article26. Park BS, Kang D, Kim KK, Jeong B, Lee TH, Park JW, et al. Hypothalamic TTF-1 orchestrates the sensitivity of leptin. Mol Metab. 2022; 66:101636.
Article27. Lu K, Chen X, Yan J, Li X, Huang C, Wan Q, et al. The effect of feeding behavior on hypothalamus in obese type 2 diabetic rats with glucagon-like peptide-1 receptor agonist intervention. Obes Facts. 2018; 11:181–94.
Article28. Brown E, Wilding JP, Barber TM, Alam U, Cuthbertson DJ. Weight loss variability with SGLT2 inhibitors and GLP-1 receptor agonists in type 2 diabetes mellitus and obesity: mechanistic possibilities. Obes Rev. 2019; 20:816–28.
Article29. Bentsen MA, Mirzadeh Z, Schwartz MW. Revisiting how the brain senses glucose-and why. Cell Metab. 2019; 29:11–7.
Article30. Yoon NA, Diano S. Hypothalamic glucose-sensing mechanisms. Diabetologia. 2021; 64:985–93.
Article31. Routh VH, Hao L, Santiago AM, Sheng Z, Zhou C. Hypothalamic glucose sensing: making ends meet. Front Syst Neurosci. 2014; 8:236.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Altered Metabolic Phenotypes and Hypothalamic Neuronal Activity Triggered by Sodium-Glucose Cotransporter 2 Inhibition (Diabetes Metab J 2023;47:784-95)
- Altered Metabolic Phenotypes and Hypothalamic Neuronal Activity Triggered by Sodium-Glucose Cotransporter 2 Inhibition (Diabetes Metab J 2023;47:784-95)
- SGLT2 Inhibitors and Ketoacidosis: Pathophysiology and Management
- Sodium-Glucose Cotransporter 2 Inhibitors: Mechanisms of Action and Various Effects
- Glucose Lowering Effect of SGLT2 Inhibitors: A Review of Clinical Studies
