Overexpression of p21 Has Inhibitory Effect on Human Hematopoiesis by Blocking Generation of CD43+ Cells via Cell-Cycle Regulation
- Affiliations
-
- 1Research Center for Stem Cell Therapies, Institute of Blood Transfusion, Chinese Academy of Medical Sciences & Peking Union Medical College (CAMS & PUMC), Chengdu, China
- 2State Key Laboratory of Biotherapy, Sichuan University, Chengdu, China
- 3State Key Laboratory of Experimental Hematology, CAMS & PUMC, Tianjin, China
- KMID: 2504333
- DOI: http://doi.org/10.15283/ijsc20033
Abstract
- Background and Objectives
p21, an important member of the Cip/Kip family, is involved in inhibitory effects of RUNX1b overexpression during the early stage of human hematopoiesis.
Methods and Results
We established a human embryonic stem cell (hESC) line with inducible expression of p21 (p21/hESCs). Overexpression of p21 did not influence either mesoderm induction or emergence of CD34+ cells, but it significantly decreased the production of CD43+ cells and changed the expression profile of hematopoiesis-related factors, leading to the negative effects of p21 on hematopoiesis.
Conclusions
In RUNX1b/hESC co-cultures when RUNX1b was induced from D0, perturbation of the cell cycle caused by upregulation of p21 probably prevented the appearance of CD43+ cells, but not CD34+ cells. The mechanisms via which CD34+ cells are blocked by RUNX1b overexpression remain to be elucidated.
Keyword
Figure
-

Fig. 1 p21 is involved in inhibitory effects of RUNX1b on the mesoderm-hemogenesis transition. D0-induced RUNX1b/hESC co-cultures with AGM-S3 cells at D4, (A) Cell-cycle analyses revealed that the proportion of cells in S phase dropped significantly when RUNX1b was overexpressed from D0, and that these inhibitory effects could be partially rescued by RepSox. (B, C) qRT-PCR and western blotting analysis showed that p21 was upregulated when DOX was added from D0, and that this effect could be counteracted by 0.33 μM RepSox. Grayscale scanning analyses were performed using Gel-Pro Analyzer 4.
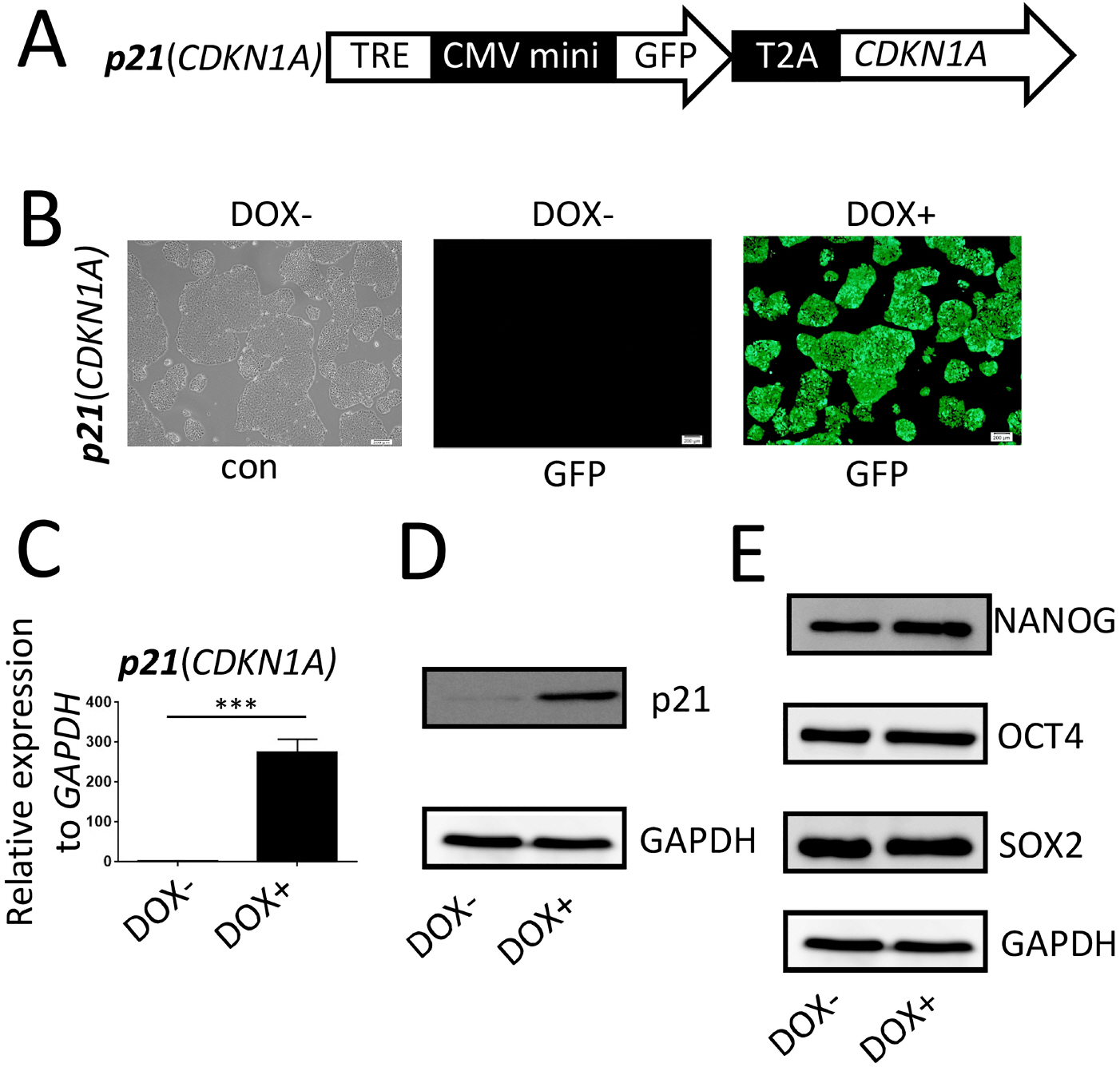
Fig. 2 Construction and confirmation of inducible p21/hESC lines. (A) Schematic representation of the PiggyBac constructs used to inducibly express p21. TRE, tet-on regulation element; CMV mini, cytomegalovirus minimum promoter; T2A, Thosea asigna virus 2A peptide. (B) After p21/hESCs were induced for 48 h, the cells were imaged by fluorescence microscopy to observe co-expression of GFP. (C, D) qRT-PCR and western blotting were used to confirm that inducible expression of p21 was highly stringent and efficient at the transcriptional and protein levels. (E) Pluripotency of p21/hESCs (non-induced or induced) was confirmed by western blotting for SOX2, OCT4, and NANOG.
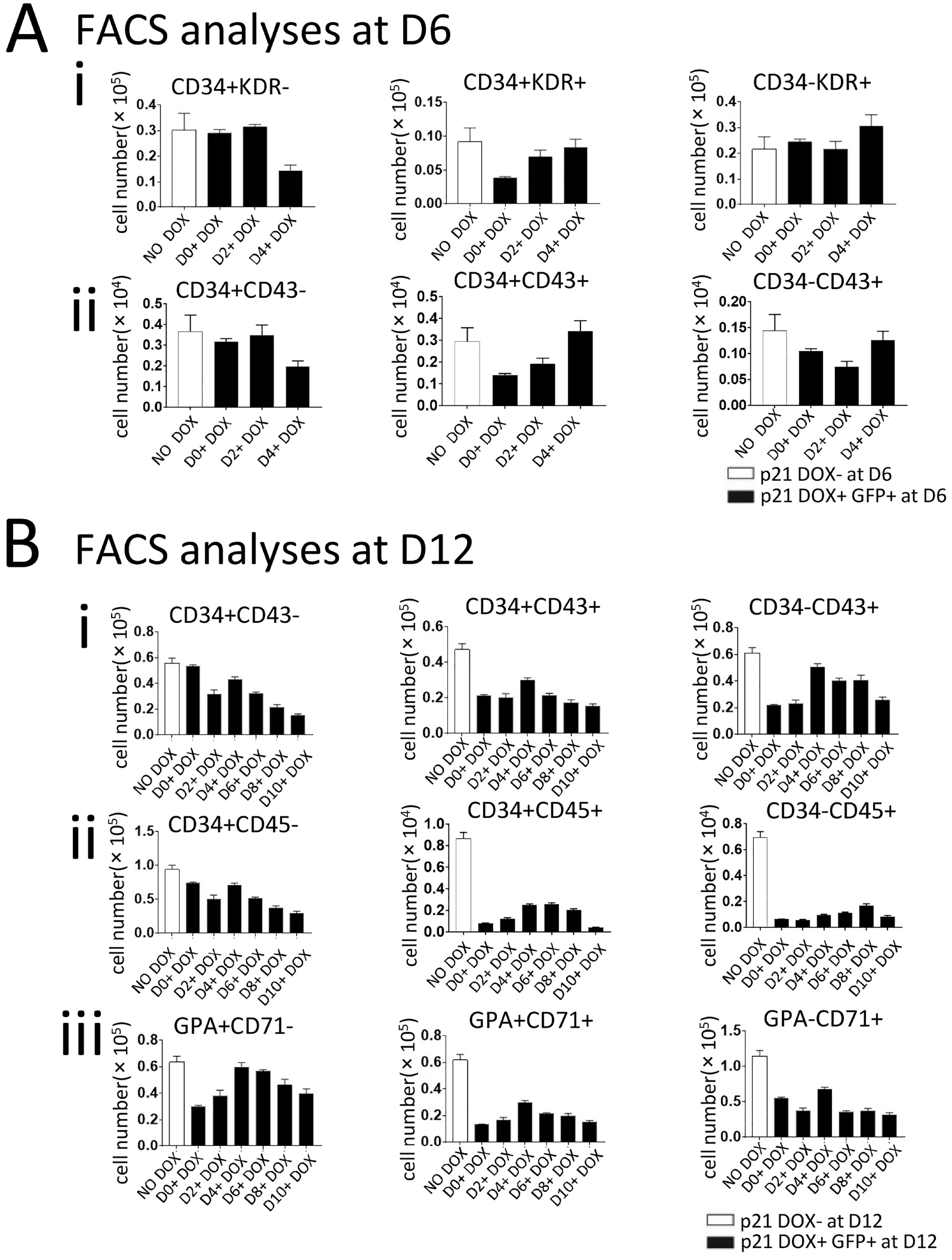
Fig. 3 Overexpression of p21 blocks hematopoiesis in co-culture with AGM-S3 cells. p21/hESC co-cultures with AGM-S3 cells were treated with DOX from D0, D2, D4, D6, D8, or D10, and then subjected to FACS with antibodies against CD34/KDR and CD34/CD43 (at D6) or GPA/CD71, CD34/CD43, and CD34/CD45 (at D12) to compare non-induced co-cultures and the GFP+ fraction of co-cultures treated with DOX. (A) When p21 was overexpressed from the early stage, the abundance of D34+KDR− and CD34+CD43− cells was not influenced at D6, whereas the emergence of CD34+KDR+ and CD34+CD43+ cells was significantly blocked. (B) Most hematopoietic populations, such as CD34−CD43+, CD34+CD43+, CD34+CD45+, CD34−CD45+, and GPA+CD71+, dramatically decreased at D12, regardless of when induction of p21 started. This observation indicates that p21 has strong negative effects on hematopoietic cells (CD43+) but not their endothelial precursors (CD34+).

Fig. 4 Colony assays confirmed the inhibitory effects of p21 on hematopoiesis. D0-induced or non-induced co-cultures of p21/hESCs with AGM-S3 (about 3×104 cells) at D12 were subjected to colony culture assays to assess their hematopoietic potentials. (A) The number of each type of colony derived from 3×104 co-cultured cells. p<0.05 was considered significant. (B) Typical morphologies of CFU-Mix (a), CFU-E (b), BFU-E (c), and CFU-GM (d) colonies. Scale bar, 100 μm. MGG staining of cells in BFU-E colonies (e). Scale bar, 10 μm. The hematopoietic potential of p21-induced co-cultures dropped significantly.
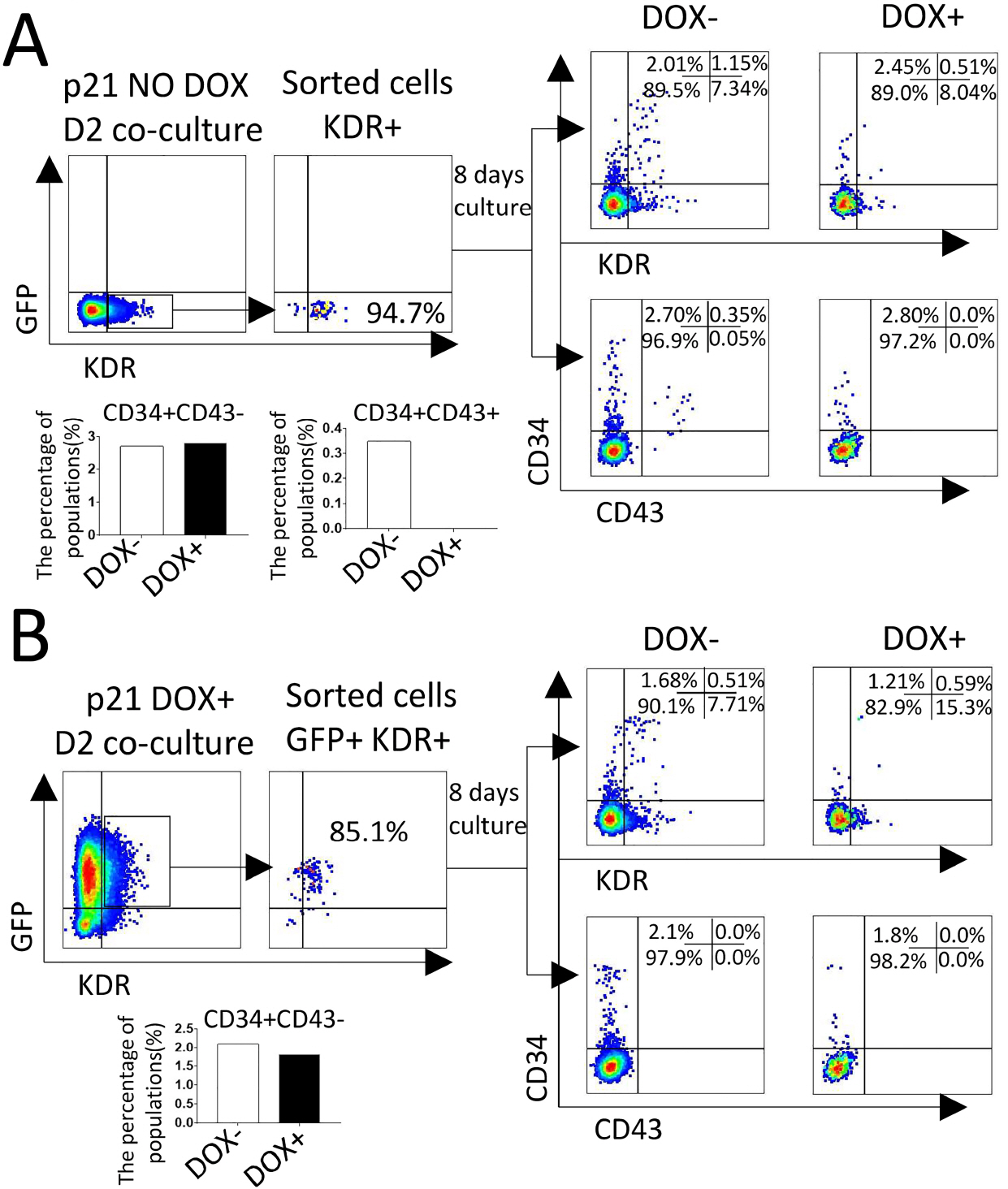
Fig. 5 Induction of p21 from the early stage severely blocked the emergence of CD43+ cells. KDR+ or GFP+KDR+ cells sorted from D0-induced or non-induced p21/hESC co-cultures at D2. About 5×103 cells were re-plated on irradiated AGM-S3 in 24-well plates or further hematopoietic culture, treated with or without DOX for 8 days, and then subjected to FACS analysis using a combination of CD34/KDR or CD34/CD43 antibodies. The results indicated that induction of p21 from D0 or D2 significantly blocked the appearance of CD43+ cells, but not CD34+ cells.
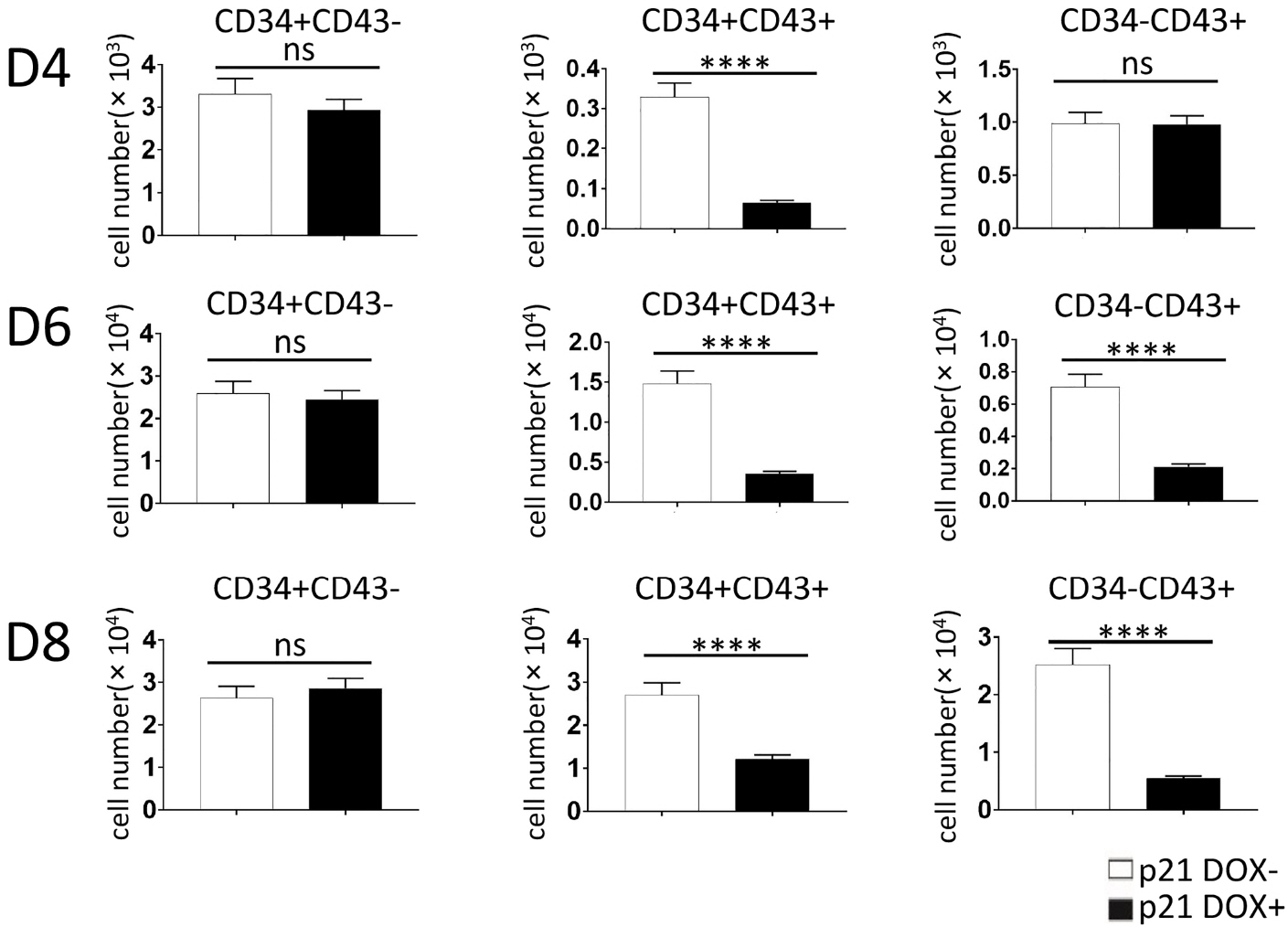
Fig. 6 Overexpression of p21 from D0 blocked the emergence of CD43+ cells in co-cultures with AGM-S3 cells. p21/hESC co-cultures were treated with DOX from D0, and then subjected to FACS analysis using combinations of anti-CD34/CD43 antibodies to compare non-induced co-cultures and the GFP+ fraction of co-cultures at D4, D6, and D8. Production of CD34+CD43− cells was not significantly influenced, whereas production of CD34−CD43+ and CD34+CD43+ dropped significantly following induction of p21, indicating that p21 has inhibitory effects on CD43+ cells but not CD34+ cells.

Fig. 7 Overexpression of p21 interfered with the expression of hematopoiesis-related genes in co-cultures with AGM-S3 cells. p21/hESCs co-cultures were treated with DOX from D0. qRT-PCR analyses at D4 revealed that hematopoiesis-related genes, including GATA1, GATA2, vWF, and LOM2, were down-regulated. Expression of CD34+ was not influenced, but RUNX1b/c was up-regulated, potentially causing the inhibitory effects of p21 on hematopoiesis.

Fig. 8 Overexpression of p21 interferes with the cell-cycle status of KDR+ and CD34+ cells in co-culture with AGM-S3 cells. In p21/hESC co-cultures with AGM-S3 cells induced from D0, cell-cycle analysis indicated that (A) the proportion of KDR+ cells in S phase dropped significantly at D4, and this effect could be counteracted by RepSox; and that (B) the proportion of CD34+ cells in G2/M phase dropped significantly at D6, but this effect could not be counteracted by RepSox.
Reference
-
References
1. Palis J, Yoder MC. 2001; Yolk-sac hematopoiesis: the first blood cells of mouse and man. Exp Hematol. 29:927–936. DOI: 10.1016/S0301-472X(01)00669-5.
Article2. Medvinsky A, Dzierzak E. 1996; Definitive hematopoiesis is autonomously initiated by the AGM region. Cell. 86:897–906. DOI: 10.1016/S0092-8674(00)80165-8.
Article3. Rhodes KE, Gekas C, Wang Y, Lux CT, Francis CS, Chan DN, Conway S, Orkin SH, Yoder MC, Mikkola HK. 2008; The emergence of hematopoietic stem cells is initiated in the placental vasculature in the absence of circulation. Cell Stem Cell. 2:252–263. DOI: 10.1016/j.stem.2008.01.001. PMID: 18371450. PMCID: PMC2888040.
Article4. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. 1998; Embryonic stem cell lines derived from human blastocysts. Science. 282:1145–1147. DOI: 10.1126/science.282.5391.1145. PMID: 9804556.
Article5. Ma F, Wang D, Hanada S, Ebihara Y, Kawasaki H, Zaike Y, Heike T, Nakahata T, Tsuji K. 2007; Novel method for efficient production of multipotential hematopoietic progenitors from human embryonic stem cells. Int J Hematol. 85:371–379. DOI: 10.1532/IJH97.06203. PMID: 17562610.
Article6. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. 2007; Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 131:861–872. DOI: 10.1016/j.cell.2007.11.019. PMID: 18035408.
Article7. Ma F, Ebihara Y, Umeda K, Sakai H, Hanada S, Zhang H, Zaike Y, Tsuchida E, Nakahata T, Nakauchi H, Tsuji K. 2008; Generation of functional erythrocytes from human embryonic stem cell-derived definitive hematopoiesis. Proc Natl Acad Sci U S A. 105:13087–13092. DOI: 10.1073/pnas.0802220105. PMID: 18755895. PMCID: PMC2526552.
Article8. Palis J. 2016; Hematopoietic stem cell-independent hematopoiesis: emergence of erythroid, megakaryocyte, and myeloid potential in the mammalian embryo. FEBS Lett. 590:3965–3974. DOI: 10.1002/1873-3468.12459. PMID: 27790707.
Article9. Han X, Liu J. 2017; Cell cycle-independent roles of p19INK4d in human terminal erythropoiesis. Chin J Cancer. 36:22. DOI: 10.1186/s40880-017-0189-4. PMID: 28228163. PMCID: PMC5322582.
Article10. Chen B, Teng J, Liu H, Pan X, Zhou Y, Huang S, Lai M, Bian G, Mao B, Sun W, Zhou Q, Yang S, Nakahata T, Ma F. 2017; Inducible overexpression of RUNX1b/c in human embryonic stem cells blocks early hematopoiesis from mesoderm. J Mol Cell Biol. 9:262–273. DOI: 10.1093/jmcb/mjx032. PMID: 28992293.
Article11. Xiong Y, Hannon GJ, Zhang H, Casso D, Kobayashi R, Beach D. 1993; p21 is a universal inhibitor of cyclin kinases. Nature. 366:701–704. DOI: 10.1038/366701a0. PMID: 8259214.
Article12. Ouellet S, Vigneault F, Lessard M, Leclerc S, Drouin R, Guérin SL. 2006; Transcriptional regulation of the cyclin-dependent kinase inhibitor 1A (p21) gene by NFI in proliferating human cells. Nucleic Acids Res. 34:6472–6487. DOI: 10.1093/nar/gkl861. PMID: 17130157. PMCID: PMC1702497.
Article13. Stier S, Cheng T, Forkert R, Lutz C, Dombkowski DM, Zhang JL, Scadden DT. 2003; Ex vivo targeting of p21Cip1/Waf1 permits relative expansion of human hematopoietic stem cells. Blood. 102:1260–1266. DOI: 10.1182/blood-2002-10-3053. PMID: 12702511.
Article14. Matsumura I, Ishikawa J, Nakajima K, Oritani K, Tomiyama Y, Miyagawa J, Kato T, Miyazaki H, Matsuzawa Y, Kanakura Y. 1997; Thrombopoietin-induced differentiation of a human megakaryoblastic leukemia cell line, CMK, involves transcriptional activation of p21(WAF1/Cip1) by STAT5. Mol Cell Biol. 17:2933–2943. DOI: 10.1128/MCB.17.5.2933. PMID: 9111365. PMCID: PMC232145.
Article15. Eilken HM, Nishikawa S, Schroeder T. 2009; Continuous single-cell imaging of blood generation from haemogenic endothelium. Nature. 457:896–900. DOI: 10.1038/nature07760. PMID: 19212410.
Article16. Mao B, Huang S, Lu X, Sun W, Zhou Y, Pan X, Yu J, Lai M, Chen B, Zhou Q, Mao S, Bian G, Zhou J, Nakahata T, Ma F. 2016; Early development of definitive erythroblasts from human pluripotent stem cells defined by expression of glycophorin A/CD235a, CD34, and CD36. Stem Cell Reports. 7:869–883. DOI: 10.1016/j.stemcr.2016.09.002. PMID: 27720903. PMCID: PMC5106477.
Article17. Chang J, Sun W, Zeng J, Xue Y, Zhang Y, Pan X, Zhou Y, Lai M, Bian G, Zhou Q, Liu J, Chen B, Guo F, Ma F. 2019; Establishment of an in vitro system based on AGM-S3 co-culture for screening traditional herbal medicines that stimulate hematopoiesis. J Ethnopharmacol. 240:111938. DOI: 10.1016/j.jep.2019.111938. PMID: 31077780.
Article18. Zhou Y, Zhang Y, Chen B, Dong Y, Zhang Y, Mao B, Pan X, Lai M, Chen Y, Bian G, Zhou Q, Nakahata T, Zhou J, Wu M, Ma F. 2019; Overexpression of GATA2 enhances development and maintenance of human embryonic stem cell-derived hematopoietic stem cell-like progenitors. Stem Cell Reports. 13:31–47. DOI: 10.1016/j.stemcr.2019.05.007. PMID: 31178416. PMCID: PMC6626852.
Article19. Palpant NJ, Wang Y, Hadland B, Zaunbrecher RJ, Redd M, Jones D, Pabon L, Jain R, Epstein J, Ruzzo WL, Zheng Y, Bernstein I, Margolin A, Murry CE. 2017; Chromatin and transcriptional analysis of mesoderm progenitor cells identifies HOPX as a regulator of primitive hematopoiesis. Cell Rep. 20:1597–1608. DOI: 10.1016/j.celrep.2017.07.067. PMID: 28813672. PMCID: PMC5576510.
Article20. Yuan X, Braunstein EM, Ye Z, Liu CF, Chen G, Zou J, Cheng L, Brodsky RA. 2013; Generation of glycosylphosphatidylinositol anchor protein-deficient blood cells from human induced pluripotent stem cells. Stem Cells Transl Med. 2:819–829. DOI: 10.5966/sctm.2013-0069. PMID: 24113066. PMCID: PMC3808197.
Article21. Vodyanik MA, Thomson JA, Slukvin II. 2006; Leukosialin (CD43) defines hematopoietic progenitors in human embryonic stem cell differentiation cultures. Blood. 108:2095–2105. DOI: 10.1182/blood-2006-02-003327. PMID: 16757688. PMCID: PMC1895535.
Article22. Gartel AL, Radhakrishnan SK. 2005; Lost in transcription: p21 repression, mechanisms, and consequences. Cancer Res. 65:3980–3985. DOI: 10.1158/0008-5472.CAN-04-3995. PMID: 15899785.
Article23. Cheng T, Rodrigues N, Shen H, Yang Y, Dombkowski D, Sykes M, Scadden DT. 2000; Hematopoietic stem cell quiescence maintained by p21cip1/waf1. Science. 287:1804–1808. DOI: 10.1126/science.287.5459.1804. PMID: 10710306.
Article24. Waga S, Hannon GJ, Beach D, Stillman B. 1994; The p21 inhibitor of cyclin-dependent kinases controls DNA replication by interaction with PCNA. Nature. 369:574–578. DOI: 10.1038/369574a0. PMID: 7911228.
Article25. Sun W, Teng J, Zeng J, Xue Y, Chang J, Zhang Y, Pan X, Zhou Y, Lai M, Bian G, Zhou Q, Liu J, Chen B, Ma F. 2019; The piggyBac-based double-inducible binary vector system: a novel universal platform for studying gene functions and interactions. Plasmid. 105:102420. DOI: 10.1016/j.plasmid.2019.102420. PMID: 31265838.
Article26. Xiao Y, Wang J, Song H, Zou P, Zhou D, Liu L. 2013; CD34+ cells from patients with myelodysplastic syndrome present different p21 dependent premature senescence. Leuk Res. 37:333–340. DOI: 10.1016/j.leukres.2012.11.006. PMID: 23219618.
Article27. Albanese P, Chagraoui J, Charon M, Cocault L, Dusanter-Fourt I, Romeo PH, Uzan G. 2002; Forced expression of p21 in GPIIb-p21 transgenic mice induces abnormalities in the proliferation of erythroid and megakaryocyte progenitors and primitive hematopoietic cells. Exp Hematol. 30:1263–1272. DOI: 10.1016/S0301-472X(02)00933-5.
Article28. Fortunel NO, Hatzfeld A, Hatzfeld JA. 2000; Transforming growth factor-beta: pleiotropic role in the regulation of hematopoiesis. Blood. 96:2022–2036. DOI: 10.1182/blood.V96.6.2022. PMID: 10979943.
Article29. Cheng T, Shen H, Rodrigues N, Stier S, Scadden DT. 2001; Transforming growth factor beta 1 mediates cell-cycle arrest of primitive hematopoietic cells independent of p21(Cip1/Waf1) or p27(Kip1). Blood. 98:3643–3649. DOI: 10.1182/blood.V98.13.3643. PMID: 11739168.
Article30. Ducos K, Panterne B, Fortunel N, Hatzfeld A, Monier MN, Hatzfeld J. 2000; p21(cip1) mRNA is controlled by endogenous transforming growth factor-beta1 in quiescent human hematopoietic stem/progenitor cells. J Cell Physiol. 184:80–85. DOI: 10.1002/(SICI)1097-4652(200007)184:1<80::AID-JCP8>3.0.CO;2-Q. PMID: 10825236.
Article31. Cordenonsi M, Dupont S, Maretto S, Insinga A, Imbriano C, Piccolo S. 2003; Links between tumor suppressors: p53 is required for TGF-beta gene responses by cooperating with Smads. Cell. 113:301–314. DOI: 10.1016/S0092-8674(03)00308-8. PMID: 12732139.32. Dutto I, Tillhon M, Cazzalini O, Stivala LA, Prosperi E. 2015; Biology of the cell cycle inhibitor p21(CDKN1A): molecular mechanisms and relevance in chemical toxicology. Arch Toxicol. 89:155–178. DOI: 10.1007/s00204-014-1430-4. PMID: 25514883.
Article33. Warbrick E, Lane DP, Glover DM, Cox LS. 1995; A small peptide inhibitor of DNA replication defines the site of interaction between the cyclin-dependent kinase inhibitor p21WAF1 and proliferating cell nuclear antigen. Curr Biol. 5:275–282. DOI: 10.1016/S0960-9822(95)00058-3.
Article34. Chen J, Saha P, Kornbluth S, Dynlacht BD, Dutta A. 1996; Cyclin-binding motifs are essential for the function of p21CIP1. Mol Cell Biol. 16:4673–4682. DOI: 10.1128/MCB.16.9.4673. PMID: 8756624. PMCID: PMC231467.
Article35. Oku T, Ikeda S, Sasaki H, Fukuda K, Morioka H, Ohtsuka E, Yoshikawa H, Tsurimoto T. 1998; Functional sites of human PCNA which interact with p21 (Cip1/Waf1), DNA polymerase delta and replication factor C. Genes Cells. 3:357–369. DOI: 10.1046/j.1365-2443.1998.00199.x. PMID: 9734782.
Article36. Gottifredi V, McKinney K, Poyurovsky MV, Prives C. 2004; Decreased p21 levels are required for efficient restart of DNA synthesis after S phase block. J Biol Chem. 279:5802–5810. DOI: 10.1074/jbc.M310373200. PMID: 14597617.
Article37. Cazzalini O, Perucca P, Riva F, Stivala LA, Bianchi L, Vannini V, Ducommun B, Prosperi E. 2003; p21CDKN1A does not interfere with loading of PCNA at DNA replication sites, but inhibits subsequent binding of DNA polymerase delta at the G1/S phase transition. Cell Cycle. 2:596–603. DOI: 10.4161/cc.2.6.502. PMID: 14504476.38. Smits VA, Klompmaker R, Vallenius T, Rijksen G, Mäkela TP, Medema RH. 2000; p21 inhibits Thr161 phosphorylation of Cdc2 to enforce the G2 DNA damage checkpoint. J Biol Chem. 275:30638–30643. DOI: 10.1074/jbc.M005437200. PMID: 10913154.
Article39. Charrier-Savournin FB, Château MT, Gire V, Sedivy J, Piette J, Dulic V. 2004; p21-Mediated nuclear retention of cyclin B1-Cdk1 in response to genotoxic stress. Mol Biol Cell. 15:3965–3976. DOI: 10.1091/mbc.e03-12-0871. PMID: 15181148. PMCID: PMC515331.
Article40. Gillis LD, Leidal AM, Hill R, Lee PW. 2009; p21Cip1/WAF1 mediates cyclin B1 degradation in response to DNA damage. Cell Cycle. 8:253–256. DOI: 10.4161/cc.8.2.7550. PMID: 19158493.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Effect of Ginseng on the Expression of the Onco-suppressor Gene p21in Human Breast-Cancer Cell Lines
- p53 gene transfer does not enhance E2F-1-mediated apoptosis in human colon cancer cells
- The Effect of Transfer of p21/WAF1 Gene on Kidney and Bladder Cancer Cell Lines
- Apoptotic effect of IP6 was not enhanced by co-treatment with myo-inositol in prostate carcinoma PC3 cells
- Effects of p27 Overexpression on Head and Neck Squamous Cell Carcinoma Cell Lines
