Participation of CCL1 in Snail-Positive Fibroblasts in Colorectal Cancer Contribute to 5-Fluorouracil/Paclitaxel Chemoresistance
- Affiliations
-
- 1Department of Microbial and Biochemical Pharmacy, School of Pharmaceutical Sciences, Sun Yat-sen University, Guangzhou, China. dujun@mail.sysu.edu.cn
- 2Markey Cancer Center, University of Kentucky, Lexington, KY, USA.
- 3Department of Obstetrics, Guangzhou Institute of Pediatrics, Guangzhou Women and Children's Medical Center, Guangzhou Medical University, Guangzhou, China.
- 4Department of Pharmacology, School of Pharmaceutical Sciences, Jinan University, Guangzhou, China.
- KMID: 2417878
- DOI: http://doi.org/10.4143/crt.2017.356
Abstract
- PURPOSE
Cancer-associated fibroblasts (CAFs) activated by cancer cells has a central role in development and malignant biological behavior in colorectal cancer (CRC). Adult fibroblasts do not express Snail, but Snail-positive fibroblasts are discovered in the stroma of malignant CRC and reported to be the key role to chemoresistance. However, the reciprocal effect of CAFs expressed Snail to chemoresistance on CRC cells and the underlying molecular mechanisms are not fully characterized.
MATERIALS AND METHODS
Snail-overexpressed 3T3 stable cell lines were generated by lipidosome and CT26 mixed with 3T3-Snail subcutaneous transplanted CRC models were established by subcutaneous injection. Cell Counting Kit-8, flow cytometry and western blotting assays were performed, and immunohistochemistry staining was studied. The cytokines participated in chemoresistance was validated with reverse transcriptase-polymerase chain reaction and heatmap.
RESULTS
Snail-expression fibroblasts are discovered in human and mouse spontaneous CRCs. Overexpression of Snail induces 3T3 fibroblasts transdifferentiation to CAFs. CT26 co-cultured with 3T3-Snail resisted the impairment from 5-fluorouracil and paclitaxel in vitro. The subcutaneous transplanted tumor models included 3T3-Snail cells develop without restrictions even after treating with 5-fluorouracil or paclitaxel. Moreover, these chemoresistant processes may be mediated by CCL1 secreted by Snail-expression fibroblasts via transforming growth factor β/nuclear factor-κB signaling pathways.
CONCLUSION
Taken together, Snail-expressing 3T3 fibroblasts display CAFs properties that support 5-fluorouracil and paclitaxel chemoresistance in CRC via participation of CCL1 and suggest that inhibition of the Snail-expression fibroblasts in tumor may be a useful strategy to limit chemoresistance.
MeSH Terms
-
Adult
Animals
Blotting, Western
Cell Count
Cell Line
Colorectal Neoplasms*
Cytokines
Drug Resistance, Multiple
Fibroblasts*
Flow Cytometry
Fluorouracil
Humans
Immunohistochemistry
In Vitro Techniques
Injections, Subcutaneous
Mice
Paclitaxel
Snails
Transforming Growth Factors
Cytokines
Fluorouracil
Paclitaxel
Transforming Growth Factors
Figure
-
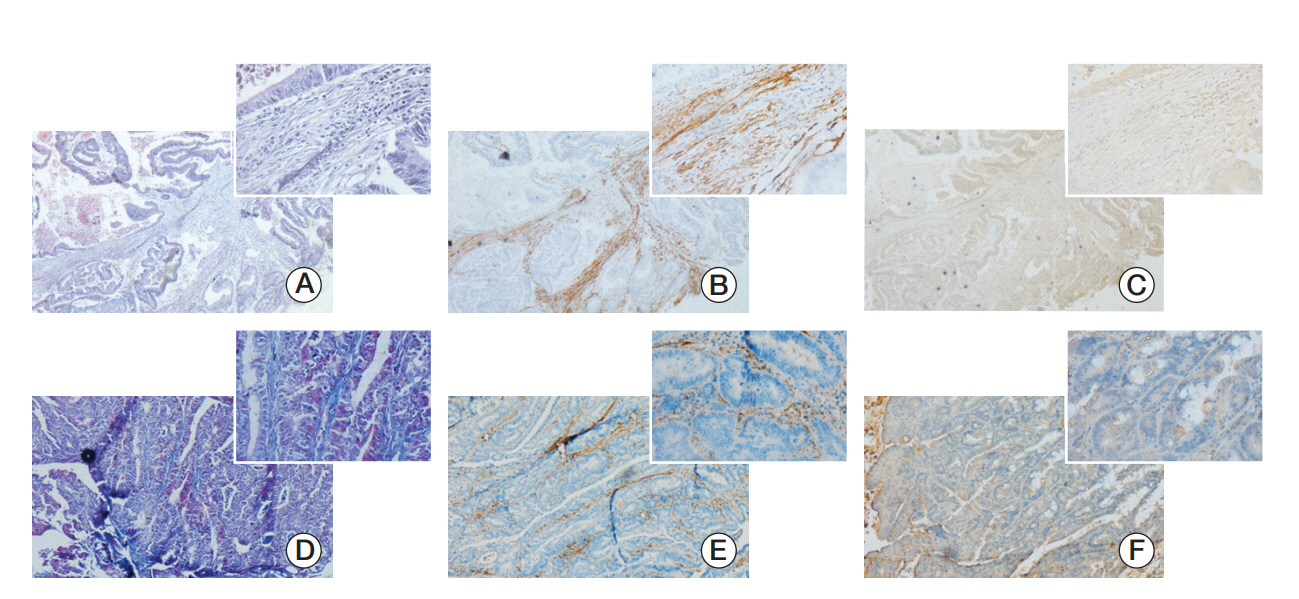
Fig. 1. Expression of Snail in cancer associated fibroblasts of human colorectal cancer tissues and Kunming (KM) mice’s spontaneous colorectal cancers. (A, D) Masson staining of paraffin-embedded colorectal cancer tissue sections (×40) and the figure in the top right corner was enlarged (×400). (B, E) Activated fibroblast marker α-smooth muscle actin expression in paraffin-embedded colorectal cancer tissue sections was detected by immunohistochemistry. (C, F) Immunohistochemistry was used to detect the Snail expression. The paraffin-embedded colorectal cancer tissue sections of panels A, B and C were from human and panels D, E and F were KM mice’s spontaneous colorectal cancers tissue sections. Besides tumor cells, cancer-associated fibroblasts expressed Snail.
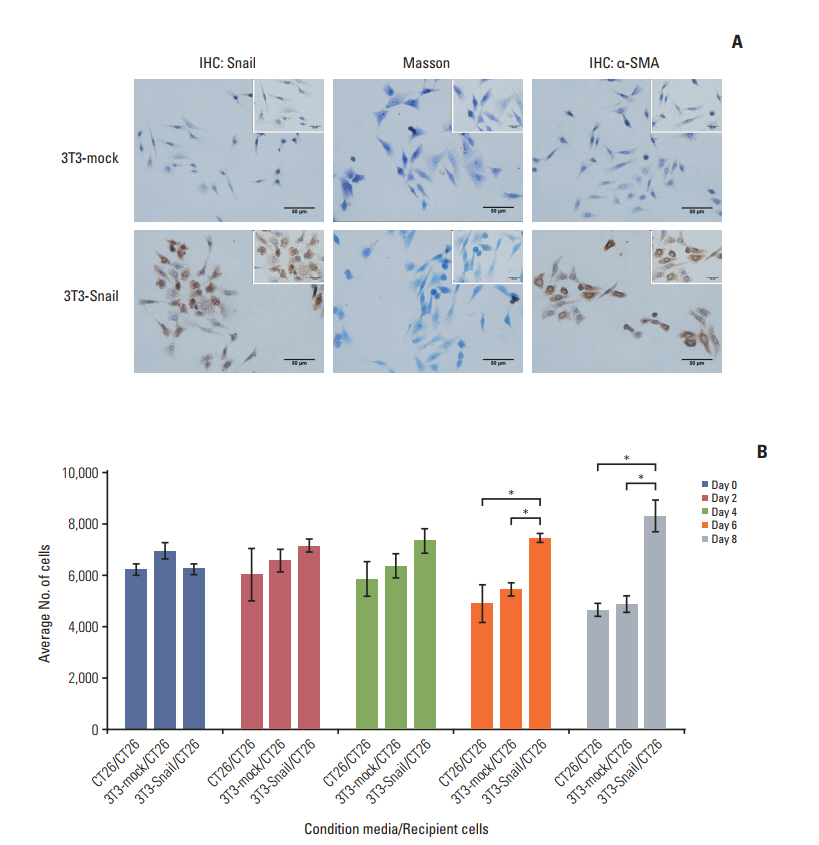
Fig. 2. Fibroblasts, overexpressed Snail, display cancer associated fibroblasts (CAFs) properties whose conditioned media ensures survival of colorectal cancer cells. (A) Snail and α-smooth muscle actin (α-SMA) expressions in 3T3-mock and 3T3- Snail stable cell lines grown on a coverslip were detected by immunohistochemistry (IHC) (×400; inset ×1,000). Masson staining of 3T3-mock and 3T3-Snail were also applied. 3T3-Snail stable cell lines expressed α-SMA, present characteristics of CAFs. (B) CT26 cells were grown in 3T3-mock‒conditioned media or 3T3-Snail‒conditioned media without fetal bovine serum for 8 days and total cells were counted daily by Cell Counting Kit 8. The conditioned media of 3T3-Snail guaranteed the survival of CT26 (*p < 0.05).
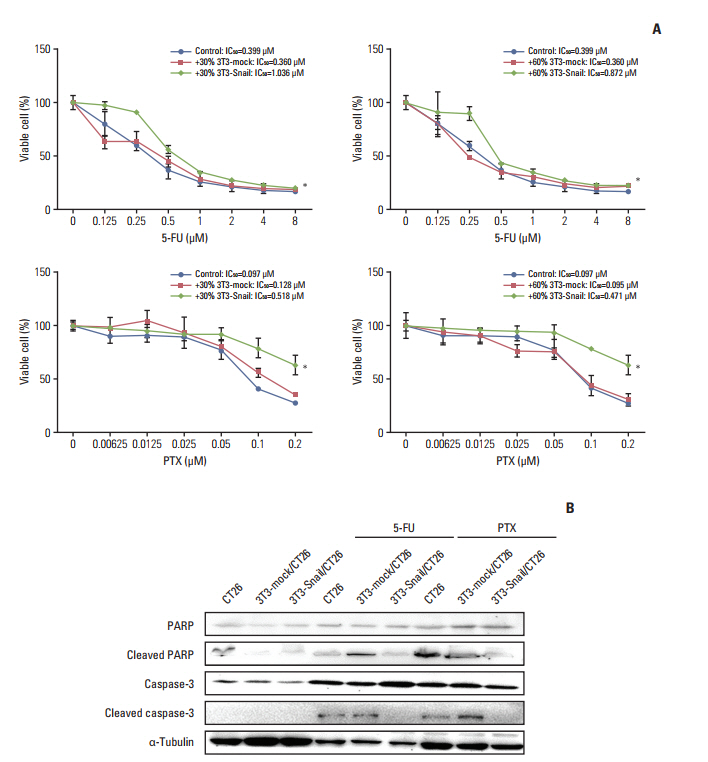
Fig. 3. Snail-overexpressed fibroblasts-conditioned media (CM) induce 5-fluorouracil (5-FU)/paclitaxel (PTX) resistance in colorectal cancer cells in vitro. (A) Murine colorectal cancer CT26 cells were treated with the indicated dose of 5-FU/PTX alone or co-cultured with 30%/60% 3T3-mock/3T3-Snail-conditioned media for 48 hours, and the cell viability was examined by Cell Counting Kit 8 assay. The IC50 values of 5-FU/PTX for CT26 were determined (*p < 0.05). (B) The levels of the poly(ADP-ribose) polymerase (PARP), cleaved PARP, caspase-3, and cleaved caspase-3 proteins from CT26 alone or co-cultured with 3T3-mock/3T3-Snail CM were analyzed by Western blotting. 3T3-Snail CM contained something induced 5-FU/PTX resistance in CT26.
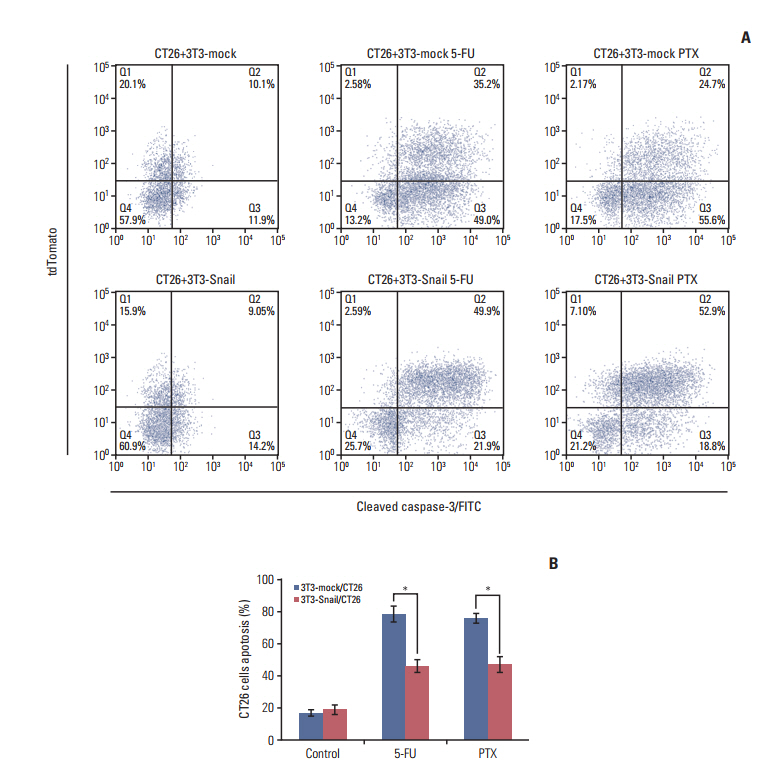
Fig. 4. Snail-overexpressed fibroblasts directly induce 5-fluorouracil (5-FU)/paclitaxel (PTX) resistance in colorectal cancer cells in vitro. CT26 were pre-cultured with 3T3-mock or 3T3-Snail by 1:2 ratios for 24 hours and further treated with 5-FU (0.8 μM)/PTX (0.2 μM) for an additional 48 hours. (A) Cells were collected and incubated with cleaved caspase-3–FITC, the percentage of cleaved caspase-3–FITC in CT26 cells was detected using flow cytometry. (B) Q3/(Q3+Q4) panel: quantifications of caspase-3 at CT26 in a representative mammary gland (*p < 0.05). 3T3-Snail decreased CT26 cells apoptosis from 5-FU/PTX.
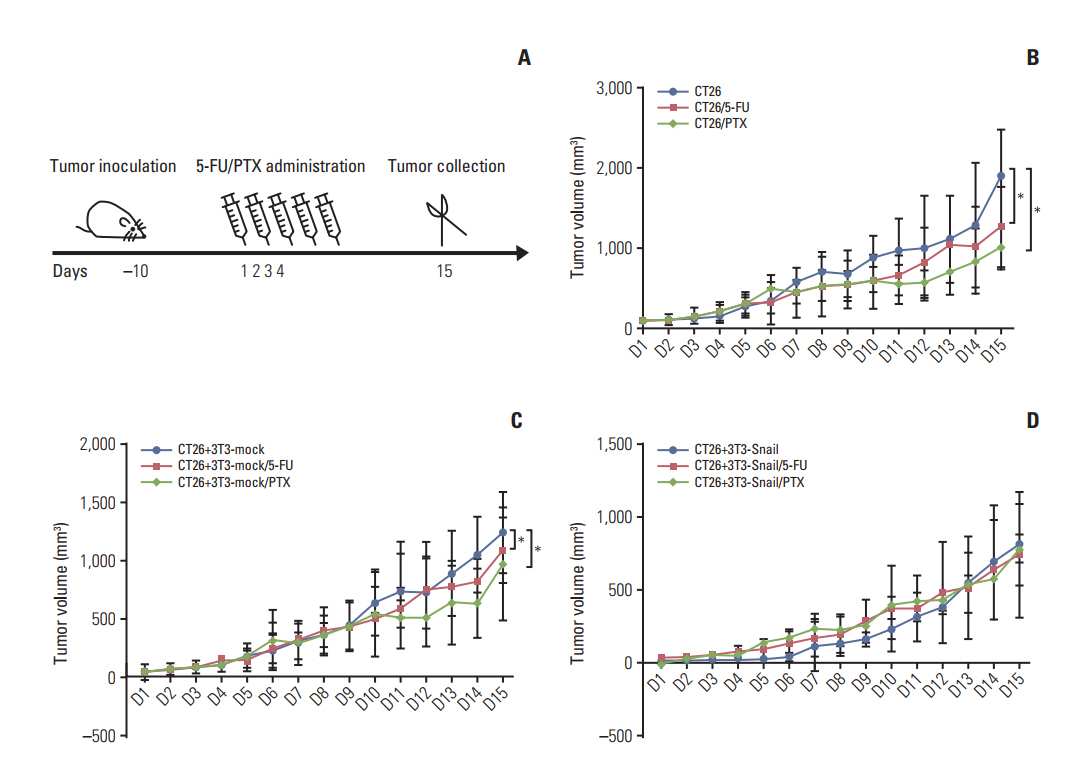
Fig. 5. Snail-overexpressed fibroblasts induce 5-fluorouracil (5-FU)/paclitaxel (PTX) resistance in the subcutaneous transplanted in BABL/c mice. (A) Treatment protocol of scheduling administration of 5-FU (30 mg/kg)/PTX (15 mg/kg) in tumorbearing mice by tail vein injection. (B, C) Tumor growth curves showed that tumors grew much slower in groups of tumor CT26 cells alone and CT26 cells mixed with 3T3-mock treated with 5-FU or PTX than the groups without treatment (n=5, *p < 0.05). (D) Meanwhile, tumor grew without any restriction in the groups of 3T3-Snail mixed with CT26 tumor cells.
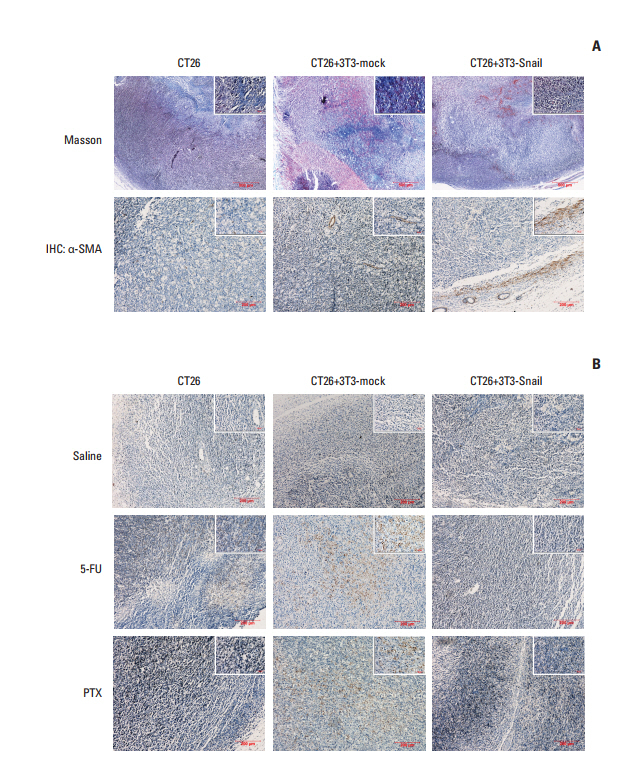
Fig. 6. Cleaved caspase-3 expression in the tumor tissue sections of subcutaneous transplanted in BALB/c mice. (A) Representative images of Masson-stained and α-smooth muscle actin (α-SMA)‒stained in paraffin sections of CT26 tumor tissue were shown. The tumor of CT26 mixed with 3T3-Snail expressed much more α-SMA than other groups. (B) Representative images of cleaved caspase-3 staining in paraffin sections of CT26 tumor tissue were shown. The expression of cleaved caspase-3 showed much less in the group of CT26 mixed with 3T3-Snail after treatment with 5-fluorouracil (5-FU)/paclitaxel (PTX) than other groups. IHC, immunohistochemistry.
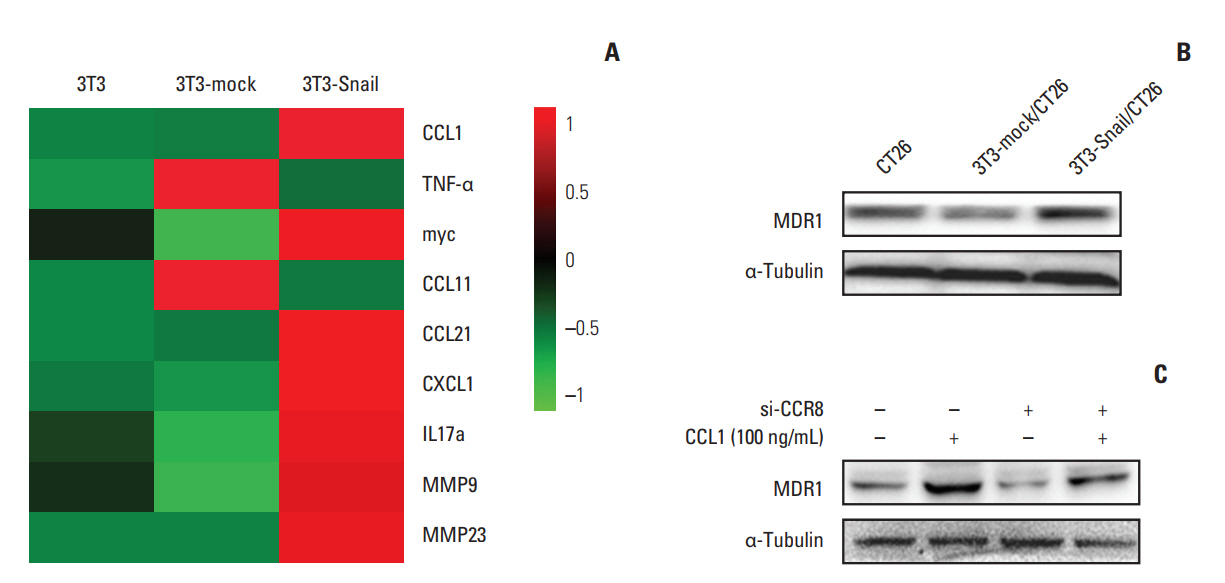
Fig. 7. Role of chemokine C-C motif ligand-1 (CCL1) in 3T3-Snail contribute to chemoresistance in colorectal cancer. (A) Heatmap representation of RNA expression levels in 3T3, 3T3-mock, and 3T3-Snail. The data was shown graphically in visualized by using a heatmap tool, OmicShare Tool. (B) The protein level of multidrug resistance protein 1 (MDR1) in CT26 after co-cultured with 3T3-Snail was shown by Western blot analysis. MDR1 expressed increasedly in CT26 co-cultured with 3T3-Snail cell lines. (C) CT26 cells were treated with CCL1 (100 ng/mL) and/or transfected si-CCR8 for 48 hours and the expression of MDR1 was detected by Western blotting. CCR8, chemokine (C-C motif) receptor 8; IL, interleukin; MMP, matrix metalloproteinase.

Fig. 8. Chemokine (C-C motif) ligand 1 (CCL1) contribute to chemoresistance in CT26 via transforming growth factor β (TGF-β) and nuclear factor кB (NF-кB) signaling pathways. (A) CT26 cells were treated with CCL1 for 5 minutes, 15 minutes, 30 minutes, and 1 hour, and the key proteins of several signaling pathways were detected via western blotting. (B) CT26 cells were pretreated with or without SB431542 (20 μM), BAY11-7081 (10 μM) for 2 hours and treated with CCL1 (100 ng/mL) for 48 hours, and then the expression of multidrug resistance protein 1 (MDR1) was analyzed by western blot. (C, D) Cells were pretreated with or without SB431542 (20 μM) (C)/BAY11-7081 (10 μM) (D) for 2 hours and treated with or without CCL1 (100 ng/mL) for 5 minutes, and TGF-β, NF-кB pathway key proteins were analyzed by western blot. GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Reference
-
References
1. Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011; 61:69–90.
Article2. Yao J, Huang A, Zheng X, Liu T, Lin Z, Zhang S, et al. 53BP1 loss induces chemoresistance of colorectal cancer cells to 5-fluorouracil by inhibiting the ATM-CHK2-P53 pathway. J Cancer Res Clin Oncol. 2017; 143:419–31.
Article3. De Wever O, Demetter P, Mareel M, Bracke M. Stromal myofibroblasts are drivers of invasive cancer growth. Int J Cancer. 2008; 123:2229–38.
Article4. Kalluri R. The biology and function of fibroblasts in cancer. Nat Rev Cancer. 2016; 16:582–98.
Article5. Piera-Velazquez S, Li Z, Jimenez SA. Role of endothelial-mesenchymal transition (EndoMT) in the pathogenesis of fibrotic disorders. Am J Pathol. 2011; 179:1074–80.
Article6. Pan R, Zhang Y, Zheng M, Zang B, Jin M. Hydroxysafflor yellow A suppresses MRC-5 cell activation induced by TGF-beta1 by blocking TGF-beta1 binding to TbetaRII. Front Pharmacol. 2017; 8:264.
Article7. Marsh T, Pietras K, McAllister SS. Fibroblasts as architects of cancer pathogenesis. Biochim Biophys Acta. 2013; 1832:1070–8.
Article8. Miller MD, Krangel MS. The human cytokine I-309 is a monocyte chemoattractant. Proc Natl Acad Sci U S A. 1992; 89:2950–4.
Article9. Das S, Sarrou E, Podgrabinska S, Cassella M, Mungamuri SK, Feirt N, et al. Tumor cell entry into the lymph node is controlled by CCL1 chemokine expressed by lymph node lymphatic sinuses. J Exp Med. 2013; 210:1509–28.
Article10. Spinetti G, Bernardini G, Camarda G, Mangoni A, Santoni A, Capogrossi MC, et al. The chemokine receptor CCR8 mediates rescue from dexamethasone-induced apoptosis via an ERKdependent pathway. J Leukoc Biol. 2003; 73:201–7.
Article11. Naber HP, Drabsch Y, Snaar-Jagalska BE, ten Dijke P, van Laar T. Snail and Slug, key regulators of TGF-beta-induced EMT, are sufficient for the induction of single-cell invasion. Biochem Biophys Res Commun. 2013; 435:58–63.12. Franci C, Gallen M, Alameda F, Baro T, Iglesias M, Virtanen I, et al. Snail1 protein in the stroma as a new putative prognosis marker for colon tumours. PLoS One. 2009; 4:e5595.
Article13. Wu Y, Zhou BP. Snail: more than EMT. Cell Adh Migr. 2010; 4:199–203.14. Liu C, Qi Y, Qiao R, Hou Y, Chan K, Li Z, et al. Detection of early primary colorectal cancer with upconversion luminescent NP-based molecular probes. Nanoscale. 2016; 8:12579–87.
Article15. Szymczak-Workman AL, Vignali KM, Vignali DA. Verification of 2A peptide cleavage. Cold Spring Harb Protoc. 2012; 2012:255–7.
Article16. Longley DB, Harkin DP, Johnston PG. 5-fluorouracil: mechanisms of action and clinical strategies. Nat Rev Cancer. 2003; 3:330–8.
Article17. Brito DA, Yang Z, Rieder CL. Microtubules do not promote mitotic slippage when the spindle assembly checkpoint cannot be satisfied. J Cell Biol. 2008; 182:623–9.
Article18. Porter AG, Janicke RU. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999; 6:99–104.
Article19. Van Snick J, Houssiau F, Proost P, Van Damme J, Renauld JC. I-309/T cell activation gene-3 chemokine protects murine T cell lymphomas against dexamethasone-induced apoptosis. J Immunol. 1996; 157:2570–6.20. Romano M, De Francesco F, Gringeri E, Giordano A, Ferraro GA, Di Domenico M, et al. Tumor microenvironment versus cancer stem cells in cholangiocarcinoma: synergistic effects? J Cell Physiol. 2016; 231:768–76.
Article21. Hu Y, Yan C, Mu L, Huang K, Li X, Tao D, et al. Fibroblastderived exosomes contribute to chemoresistance through priming cancer stem cells in colorectal cancer. PLoS One. 2015; 10:e0125625.
Article22. Lotti F, Jarrar AM, Pai RK, Hitomi M, Lathia J, Mace A, et al. Chemotherapy activates cancer-associated fibroblasts to maintain colorectal cancer-initiating cells by IL-17A. J Exp Med. 2013; 210:2851–72.
Article23. Stanisavljevic J, Loubat-Casanovas J, Herrera M, Luque T, Pena R, Lluch A, et al. Snail1-expressing fibroblasts in the tumor microenvironment display mechanical properties that support metastasis. Cancer Res. 2015; 75:284–95.
Article24. Mukaida N, Sasaki S. Fibroblasts, an inconspicuous but essential player in colon cancer development and progression. World J Gastroenterol. 2016; 22:5301–16.
Article25. Lu LL, Chen XH, Zhang G, Liu ZC, Wu N, Wang H, et al. CCL21 facilitates chemoresistance and cancer stem cell-Like properties of colorectal cancer cells through AKT/GSK-3beta/Snail signals. Oxid Med Cell Longev. 2016; 2016:5874127.26. Nunes JJ, Pandey SK, Yadav A, Goel S, Ateeq B. Targeting NF-kappa B signaling by artesunate restores sensitivity of castrate-resistant prostate cancer cells to antiandrogens. Neoplasia. 2017; 19:333–45.
Article27. Song Y, Kim JS, Choi EK, Kim J, Kim KM, Seo HR. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 2017; 8:21650–62.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Sensitization of 5-Fluorouracil-Resistant SNUC5 Colon Cancer Cells to Apoptosis by α-Mangostin
- Phase II study of 5-fluorouracil and recombinant interferon-gamma in patients with advanced colorectal cancer
- A study of the cytotoxicity of 5-fluorouracil in the cultured rat fibroblasts
- 5-fluorouracil and low dose leucovorin in advanced colorectal carcinoma
- A Phase II Trial of Paclitaxel, 5-fluorouracil (5-FU) and Cisplatin in Patients with Metastatic or Recurrent Gastric Cancer
