The Role of Microenvironment in Preserving the Potency of Adult Porcine Pulmonary Valve Stem Cells In Vitro
- Affiliations
-
- 1Department of Cardiothoracic Surgery, Division of Pediatric Cardiovascular Surgery, Stanford University, California, USA. fchalajour@gmail.com
- KMID: 2413508
- DOI: http://doi.org/10.15283/ijsc18020
Abstract
- BACKGROUND AND OBJECTIVE
The potency of tissue resident stem cells is regulated primarily by inputs from the local microenvironment. Isolation of stem cells through enzymatic digestion of tissue may affect epigenetic regulation of cell fate and performance. Here we employ a non-enzymatic method to harvest and investigate tissue resident stem cells from the adult porcine pulmonary valve.
METHODS AND RESULTS
The presence of c-Kit+ stem cells within the valve tissue was confirmed by immunohistochemistry. An in vitro culture of minced valve leaflets was developed under the standard conditions (37°C with 5% CO2). The viability of the cellular outgrowths was evaluated over the subsequent 12 weeks. Under this culture condition, we identified a population of non-adherent c-Kit+ cells and multiple cellular structures mimicking the phenotype of embryonic stem cells at different stages of development. Formation of multinucleated cells through cell fusion provided an active niche area for homing and interaction of the non-adherent c-Kit+ cells. Expression of pluripotency markers Oct-4 and Nanog was detected in the newly formed multinucleated cells but not in mature colonies. Partial cell fusion was shown by fluorescent live-cell tracking, which confirmed intercellular molecular exchange between donor and recipient cells, resulting in altered cytoplasmic protein expression by the recipient cell.
CONCLUSIONS
These results suggest a role for the microenvironment in decrypting the potential of the valve somatic stem cells in vitro. In addition, our data provide evidence for cell fusion, which may play a critical role in reversing somatic cell fate and spontaneous cellular reprogramming.
MeSH Terms
Figure
-

Fig. 1 Expression and colocalization of stem cell markers in the leaflet (A~J). IHC staining showed the presence of c-Kit+ (A; arrows) and rare Oct-4+ cells (B; arrows) within VECs. Expression of α-SMA was observed in a subset of multinucleated cells (C; arrows) below the endothelial surface of the ventricular side. The positive staining for PCNA showed the proliferation potential of VECs and VICs (D; arrows). The expression of pluripotent stem cell markers SSEA-1 (E), SSEA-4 (F), Tra-1-60 (G), and Tra-1-81 (H) was not detected in the representative tissue sections. Co-expression of c-Kit (red) and e-Nos (green) in VECs (I; arrows) was confirmed by immunofluorescent staining. In addition, a subset of c-Kit+ cells (red) was found in the subendothelial region directly adjacent to the α-SMA+ multinucleated cells (J; green). DAPI (blue) was used for counterstaining of the nuclei.

Fig. 2 Formation of diverse cellular structures from the leaflet during 12 weeks of tissue culture (A~P). Outgrowth of adherent fibroblast-like cells started a few hours after placing the leaflet tissue in the culture; these fibroblast-like cells stabilized the attachment of the leaflet tissue to the culture dish and provided a feeder layer for non-adherent cells (A). Retraction of the confluent feeder layer created a space (B) for formation of multinucleated syncytium-like structures (C) and a niche area (D). By the second week, bubble-like structures of differing sizes and shapes were detected (E). The phenotype of these structures was similar to those of oocytes, 2-cell embryos, 3-cell embryos (F), 4-cell embryos (G), and morulae (H). Formation of large dense colonies and hollow spheres occurred in the confluent cultures between week 3 and 12 (I). The dense colonies were formed either by recruiting the non-adherent cells to the morula-like structures (J) or by the retraction of the mature niche area (K). Interconnecting bundles were generated between adjacent niche syncytia (L, arrow), a process that often was followed by unification of the two structures to form a larger colony. Phalloidin staining (green) and DAPI nuclear counterstaining (blue) showed the structure of the embryonic-like cells and colonies. Oocyte-like cells exhibited a ring-shaped structure with an elongated and peripheral nucleus (M). Multiple nuclei were located on the periphery of 2-cell-embryo-like structure (N), indicating the possibility that other non-adherent cells would be arranged around these structures. Two-dimensional imaging of one blastula-like structure showed a round aggregation of cells with distinct borders between the cells (O). The unification of two adjacent niche areas was promoted by cytoplasmic fusion (P).
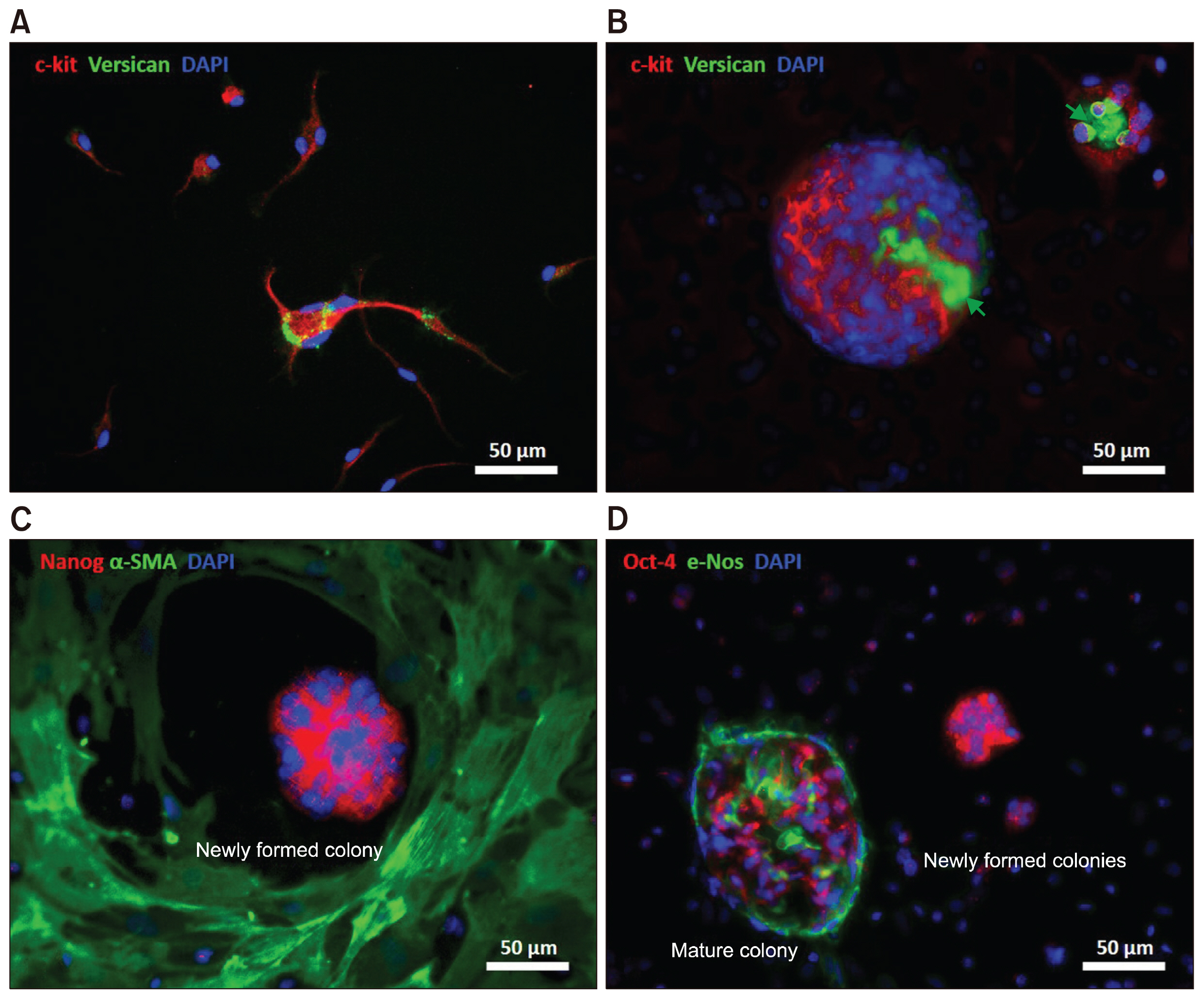
Fig. 3 Characterization of different types of multinucleated cells in a culture of the leaflet (A~D). Expression of c-Kit was detected in the majority of non-adherent mononuclear and multinuclear cells (A, B; red). Versican was expressed by multinucleated cells and found in the inner cavity of blastula-like colonies, suggesting a potential role for versican in cell aggregation and tissue regeneration (A, B; green arrows). Myofibroblast characteristics of the adherent cells surrounding the niche area were identified by expression of α-SMA (C; green). Expression of pluripotent markers Nanog and Oct-4 were detected in the newly formed niche areas before progressively decreasing during development of multinucleated colonies (C, D; red). Expression of the endothelial cell marker e-Nos in mature colonies showed spontaneous differentiation of pluripotent stem cells into endothelial cells (D; green).

Fig. 4 Tracing fusion of non-adherent cells using fluorescent dyes of two different colors (A~P). Non-adherent cells were collected and stained (separately) with green or red cell-tracker fluorescent dyes; cells of each color were returned to the originating culture. During the first few days, some of the labeled cells attached to the dish, whereas some other cells remained non-adherent (A~D). Cytoplasmic fusion of green and red labeled cells (yellow in the overlay) was observed in syncytium-like structures (E~H), bubble-like structures (I~L), and multinucleated niche areas (M~P). Migration of labeled cells into the existing and newly formed niche areas was detected (H, L, P; arrows).
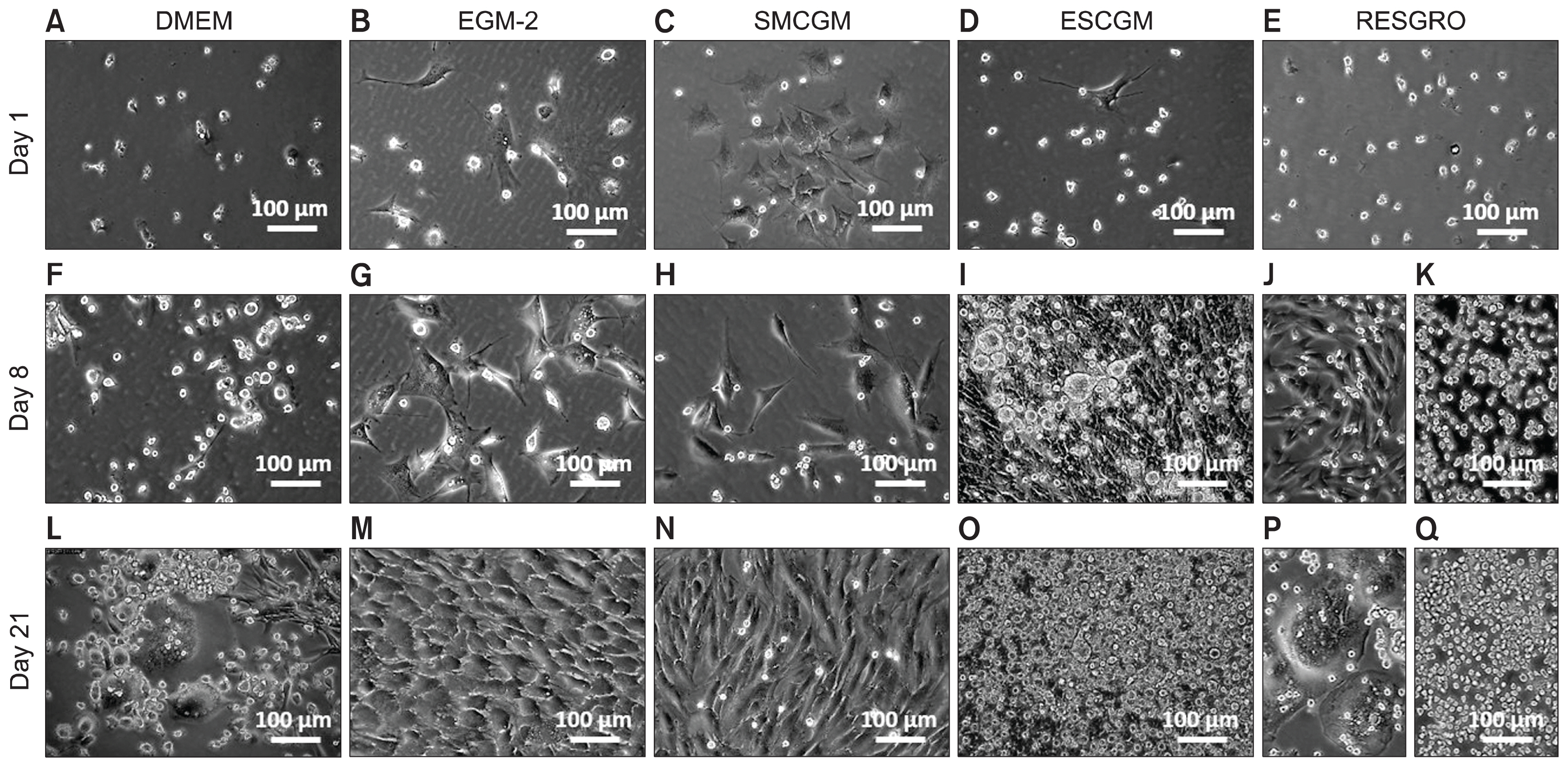
Fig. 5 Differentiation of the non-adherent cells in growth culture media. Non-adherent cells were collected from the primary culture of leaflets and grown in DMEM (A), EGM-2 (B), SMCGM (C), MSCGM (D), or RESGRO (E) medium. During the first week, non-adherent cells exhibited low levels of proliferation in DMEM medium (F) and changes in phenotype (e.g., by adhering to culture dishes) in EGM-2 or SMCGM medium (G, H). The proliferation and adhesion rates were strongly increased in MSCGM (I) or RESGRO (J, K) medium. Formation of multinucleated colonies was widely detected in DMEM or RESGRO medium (L, P) after 3 weeks, whereas proliferation of non-adherent cells and the maturation of the colonies was the evident in MSCGM or RESGRO medium (O, Q). Differentiation of non-adherent cells into cells with endothelial or myofibroblast phenotype was detected in EGM-2 or SMCGM culture, respectively (M, N).
Reference
-
References
1. Jones DL, Wagers AJ. No place like home: anatomy and function of the stem cell niche. Nat Rev Mol Cell Biol. 2008; 9:11–21. DOI: 10.1038/nrm2319.
Article2. Li M, Belmonte JC. Ground rules of the pluripotency gene regulatory network. Nat Rev Genet. 2017; 18:180–191. DOI: 10.1038/nrg.2016.156. PMID: 28045100.
Article3. Avgustinova A, Benitah SA. Epigenetic control of adult stem cell function. Nat Rev Mol Cell Biol. 2016; 17:643–658. DOI: 10.1038/nrm.2016.76. PMID: 27405257.
Article4. Jiang Y, Vaessen B, Lenvik T, Blackstad M, Reyes M, Verfaillie CM. Multipotent progenitor cells can be isolated from postnatal murine bone marrow, muscle, and brain. Exp Hematol. 2002; 30:896–904. DOI: 10.1016/S0301-472X(02)00869-X. PMID: 12160841.
Article5. Dubois SG, Floyd EZ, Zvonic S, Kilroy G, Wu X, Carling S, Halvorsen YD, Ravussin E, Gimble JM. Isolation of human adipose-derived stem cells from biopsies and liposuction specimens. Methods Mol Biol. 2008; 449:69–79. PMID: 18370084.
Article6. Smith RR, Barile L, Cho HC, Leppo MK, Hare JM, Messina E, Giacomello A, Abraham MR, Marbán E. Regenerative potential of cardiosphere-derived cells expanded from percutaneous endomyocardial biopsy specimens. Circulation. 2007; 115:896–908. DOI: 10.1161/CIRCULATIONAHA.106.655209. PMID: 17283259.
Article7. Mitchell KJ, Pannérec A, Cadot B, Parlakian A, Besson V, Gomes ER, Marazzi G, Sassoon DA. Identification and characterization of a non-satellite cell muscle resident progenitor during postnatal development. Nat Cell Biol. 2010; 12:257–266. PMID: 20118923.
Article8. Lewis FC, Henning BJ, Marazzi G, Sassoon D, Ellison GM, Nadal-Ginard B. Porcine skeletal muscle-derived multipotent PW1pos/Pax7neg interstitial cells: isolation, characterization, and long-term culture. Stem Cells Transl Med. 2014; 3:702–712. DOI: 10.5966/sctm.2013-0174. PMID: 24744394. PMCID: 4039454.
Article9. Toma JG, Akhavan M, Fernandes KJ, Barnabé-Heider F, Sadikot A, Kaplan DR, Miller FD. Isolation of multipotent adult stem cells from the dermis of mammalian skin. Nat Cell Biol. 2001; 3:778–784. DOI: 10.1038/ncb0901-778.
Article10. Paruchuri S, Yang JH, Aikawa E, Melero-Martin JM, Khan ZA, Loukogeorgakis S, Schoen FJ, Bischoff J. Human pulmonary valve progenitor cells exhibit endothelial/mesenchymal plasticity in response to vascular endothelial growth factor-A and transforming growth factor-beta2. Circ Res. 2006; 99:861–869. DOI: 10.1161/01.RES.0000245188.41002.2c. PMID: 16973908. PMCID: 2810464.
Article11. Bischoff J, Aikawa E. Progenitor cells confer plasticity to cardiac valve endothelium. J Cardiovasc Transl Res. 2011; 4:710–719. DOI: 10.1007/s12265-011-9312-0. PMID: 21789724.
Article12. Tauskela JS, Hewitt K, Kang LP, Comas T, Gendron T, Hakim A, Hogan M, Durkin J, Morley P. Evaluation of glutathione-sensitive fluorescent dyes in cortical culture. Glia. 2000; 30:329–341. DOI: 10.1002/(SICI)1098-1136(200006)30:4<329::AID-GLIA20>3.0.CO;2-R. PMID: 10797613.
Article13. Sebastià J, Cristòfol R, Martín M, Rodríguez-Farré E, Sanfeliu C. Evaluation of fluorescent dyes for measuring intracellular glutathione content in primary cultures of human neurons and neuroblastoma SH-SY5Y. Cytometry A. 2003; 51:16–25. DOI: 10.1002/cyto.a.10003.
Article14. Rabkin-Aikawa E, Mayer JE Jr, Schoen FJ. Heart valve regeneration. Adv Biochem Eng Biotechnol. 2005; 94:141–179. PMID: 15915872.
Article15. Liu AC, Joag VR, Gotlieb AI. The emerging role of valve interstitial cell phenotypes in regulating heart valve pathobiology. Am J Pathol. 2007; 171:1407–1418. DOI: 10.2353/ajpath.2007.070251. PMID: 17823281. PMCID: 2043503.
Article16. Mizuno N, Kosaka M. Novel variants of Oct-3/4 gene expressed in mouse somatic cells. J Biol Chem. 2008; 283:30997–31004. DOI: 10.1074/jbc.M802992200. PMID: 18765667. PMCID: 2662171.
Article17. Lengner CJ, Welstead GG, Jaenisch R. The pluripotency regulator Oct4: a role in somatic stem cells? Cell Cycle. 2008; 7:725–728. DOI: 10.4161/cc.7.6.5573. PMID: 18239456.
Article18. Seymour T, Twigger AJ, Kakulas F. Pluripotency genes and their functions in the normal and aberrant breast and brain. Int J Mol Sci. 2015; 16:27288–27301. DOI: 10.3390/ijms161126024. PMID: 26580604. PMCID: 4661882.
Article19. Selden C, Chalmers SA, Jones C, Standish R, Quaglia A, Rolando N, Burroughs AK, Rolles K, Dhillon A, Hodgson HJ. Epithelial colonies cultured from human explanted liver in subacute hepatic failure exhibit hepatocyte, biliary epithelial, and stem cell phenotypic markers. Stem Cells. 2003; 21:624–631. DOI: 10.1634/stemcells.21-6-624. PMID: 14595121.
Article20. Zeng L, Rahrmann E, Hu Q, Lund T, Sandquist L, Felten M, O’Brien TD, Zhang J, Verfaillie C. Multipotent adult progenitor cells from swine bone marrow. Stem Cells. 2006; 24:2355–2366. DOI: 10.1634/stemcells.2005-0551. PMID: 16931778.
Article21. Gwak JM, Kim M, Kim HJ, Jang MH, Park SY. Expression of embryonal stem cell transcription factors in breast cancer: Oct4 as an indicator for poor clinical outcome and tamoxifen resistance. Oncotarget. 2017; 8:36305–36318. DOI: 10.18632/oncotarget.16750. PMID: 28422735. PMCID: 5482656.
Article22. Villodre ES, Kipper FC, Pereira MB, Lenz G. Roles of OCT4 in tumorigenesis, cancer therapy resistance and prognosis. Cancer Treat Rev. 2016; 51:1–9. DOI: 10.1016/j.ctrv.2016.10.003. PMID: 27788386.
Article23. Müller M, Hermann PC, Liebau S, Weidgang C, Seufferlein T, Kleger A, Perkhofer L. The role of pluripotency factors to drive stemness in gastrointestinal cancer. Stem Cell Res. 2016; 16:349–357. DOI: 10.1016/j.scr.2016.02.005. PMID: 26896855.
Article24. Redshaw Z, Strain AJ. Human haematopoietic stem cells express Oct4 pseudogenes and lack the ability to initiate Oct4 promoter-driven gene expression. J Negat Results Biomed. 2010; 9:2. DOI: 10.1186/1477-5751-9-2. PMID: 20356403. PMCID: 2853495.
Article25. Liedtke S, Enczmann J, Waclawczyk S, Wernet P, Kögler G. Oct4 and its pseudogenes confuse stem cell research. Cell Stem Cell. 2007; 1:364–366. DOI: 10.1016/j.stem.2007.09.003.
Article26. Lengner CJ, Camargo FD, Hochedlinger K, Welstead GG, Zaidi S, Gokhale S, Scholer HR, Tomilin A, Jaenisch R. Oct4 expression is not required for mouse somatic stem cell self-renewal. Cell Stem Cell. 2007; 1:403–415. DOI: 10.1016/j.stem.2007.07.020. PMID: 18159219. PMCID: 2151746.
Article27. Dyce PW, Wen L, Li J. In vitro germline potential of stem cells derived from fetal porcine skin. Nat Cell Biol. 2006; 8:384–390. DOI: 10.1038/ncb1388. PMID: 16565707.
Article28. Ma Z, Hu Y, Jiang G, Hou J, Liu R, Lu Y, Liu C. Spontaneous generation of germline characteristics in mouse fibrosarcoma cells. Sci Rep. 2012; 2:743. DOI: 10.1038/srep00743. PMID: 23077727. PMCID: 3473365.
Article29. Dyce PW, Shen W, Huynh E, Shao H, Villagómez DA, Kidder GM, King WA, Li J. Analysis of oocyte-like cells differentiated from porcine fetal skin-derived stem cells. Stem Cells Dev. 2011; 20:809–819. DOI: 10.1089/scd.2010.0395.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Glutathione Dynamics in the Tumor Microenvironment: A Potential Target of Cancer Stem Cells and T Cells
- Concise Review: Differentiation of Human Adult Stem Cells Into Hepatocyte-like Cells In vitro
- Bioreactor Conditioning of Valve Scaffolds Seeded Internally with Adult Stem Cells
- Endogenous Stem Cells in the Ear
- Measurement of Porcine Aortic and Pulmonary Valve Geometry and Design for Implantable Tissue Valve
