Induction of Interleukin-22 (IL-22) production in CD4+ T Cells by IL-17A Secreted from CpG-Stimulated Keratinocytes
- Affiliations
-
- 1Department of Dermatology, Chungnam National University School of Medicine, Daejeon, Korea. jhoon@cnu.ac.kr
- KMID: 2382882
- DOI: http://doi.org/10.5021/ad.2016.28.5.579
Abstract
- BACKGROUND
Interleukin-17A (IL-17A) is mainly secreted from Th17 cells that are activated by various stimuli including CpG oligodeoxynucleotide, a Toll-like receptor 9 (TLR9) ligand. Recently, it has been demonstrated that keratinocytes play an important role in the pathogenesis of psoriasis.
OBJECTIVE
To investigate the potential role of keratinocytes, we examined whether TLR9 ligand CpG induces IL-17A expression in keratinocytes.
METHODS
We used HaCaT keratinocytes as a model system, and determined CpG-induced IL-17A using enzyme-linked immunosorbent assay and Western blot.
RESULTS
When HaCaT keratinocytes were treated with CpG, the expression of several cytokines including IL-17A, tumor necrosis factor-α and CCL20 was markedly increased. Treatment with nuclear factor (NF)-κB inhibitor significantly blocked the CpG-induced IL-17A production, indicating that CpG induced IL-17A expression through the NF-κB signaling pathway. In addition, IL-17A secreted from keratinocytes stimulated the CD4⺠T cells, resulting in strong induction of IL-22 production.
CONCLUSION
Since IL-22 is an important mediator for psoriatic inflammation, our data suggest that keratinocytes can participate in the pathogenesis of psoriasis via the TLR9-dependent IL-17A production.
Keyword
MeSH Terms
Figure
-
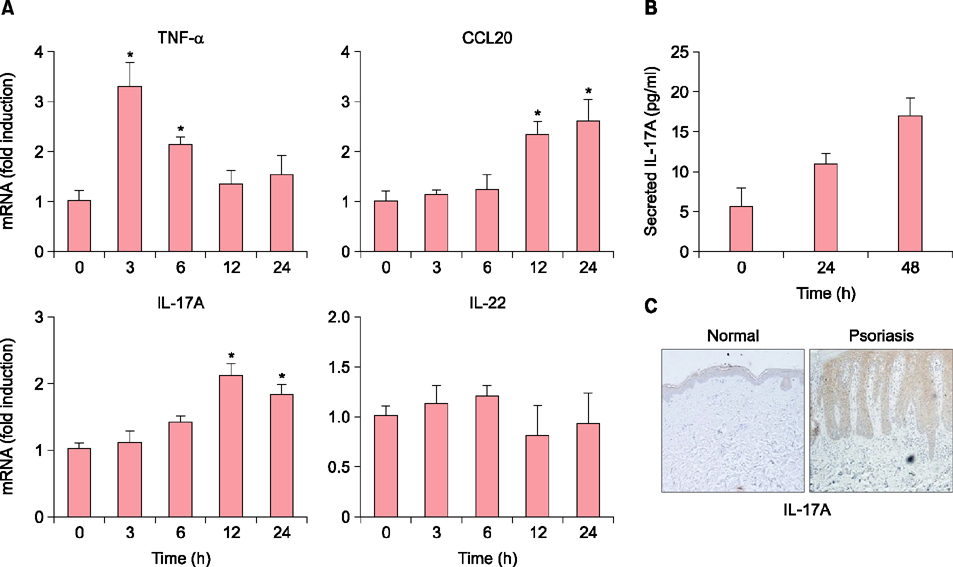
Fig. 1 CpG induces interleukin-17A (IL-17A) expression in HaCaT keratinocytes. (A) Cells were treated with 10 µM CpG for the indicated time points, and cytokine expression was assessed by quantitative real-time polymerase chain reaction. (B) After treatment with CpG, medium was collected and IL-17A was determined by ELISA. The mean values±standard deviation are averages of triplicate measurements. *p<0.05 versus time 0 hour. (C) Expression of IL-17A was detected in normal and psoriatic lesional skin tissues by immunohistochemistry. It clearly shows that expression of IL-17A is increased in psoriatic lesional skin compared to normal skin. TNF-α: tumor necrosis factor-α.
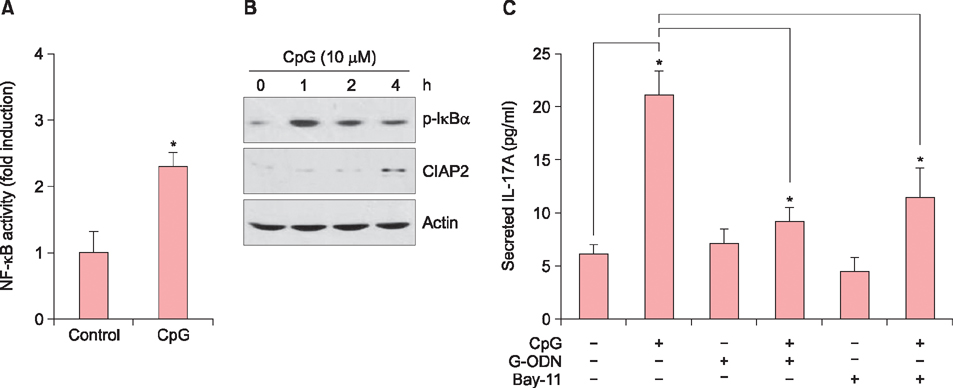
Fig. 2 CpG induces interleukin-17A (IL-17A) expression via nuclear factor (NF)-κB-dependent way. (A) HaCaT keratinocytes were transduced with 1 multiplicity of infection of NF-κB reporter adenovirus for overnight. Cells were replenished with fresh medium, and then treated with 10 µM CpG for 24 hours. Luciferase assay was performed and data are expressed as fold induction. Data represent the mean±standard deviation (SD) (n=3). *p<0.05. (B) Cells were treated with 10 µM CpG for the indicated time points, then phosphorylated-IκBα (p-IκBα) and CIAP2 were analyzed by Western blot. (C) Cells were pretreated with NF-κB inhibitor Bay-11 (10 ng/ml) or CpG antagonist G-ODN (4 µM) for 30 minutes, then treated with 10 µM CpG for 2 days. Secreted IL-17A was determined by ELISA. The mean values±SD are averages of triplicate measurements. *p<0.05.
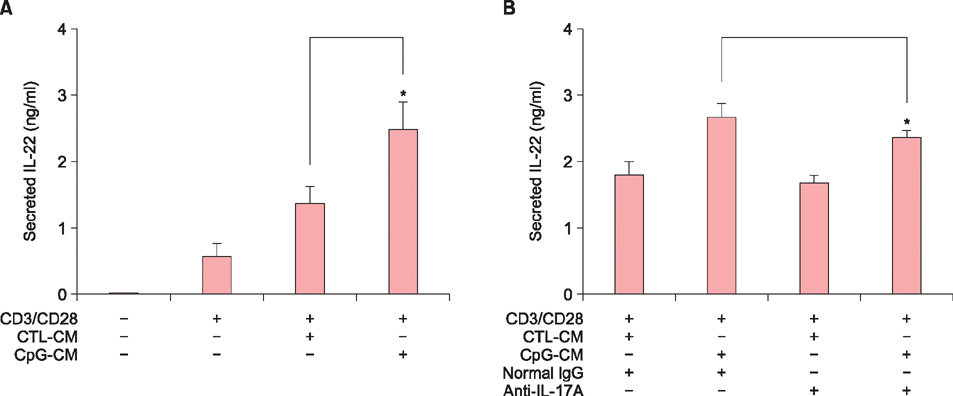
Fig. 3 Interleukin-17A (IL-17A) secreted from keratinocytes affects IL-22 production in CD4+ T cells. (A) CD4+ T cells were isolated from human blood. After a first round of stimulation with CD3/CD28, CD4+ T cells were treated with conditioned medium obtained from CpG-treated HaCaT keratinocytes (CpG-CM) and/or conditioned medium obtained from CpG-non-treated HaCaT keratinocytes (CTL-CM). For preparation of conditioned medium, HaCaT keratinocytes were treated with 10 µM CpG for 24 hours, then cells were replenished with fresh medium and incubated for a further 24 hours. After collecting the medium, cell debris was discarded by centrifugation and supernatant was used as the conditioned medium. (B) Conditioned medium was neutralized with preincubation with anti-IL-17A antibody. Normal IgG was used as a negative control. Secreted IL-22 was determined by ELISA. The mean values±standard deviation are averages of triplicate measurements. *p<0.05.
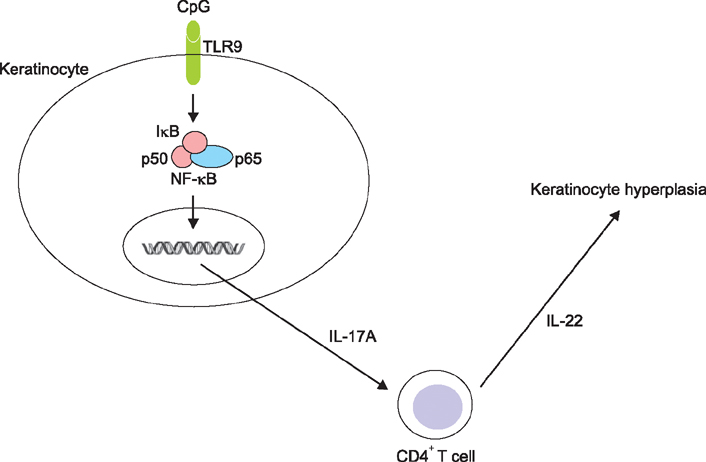
Fig. 4 Proposed model for the crosstalk between keratinocytes and CD4+ T cells. Keratinocytes recognize the Toll-like receptor 9 (TLR9) agonist and then release interleukin-17A (IL-17A) in a nuclear factor (NF)-κB-dependent way. Secreted IL-17A activates CD4+ T cells to produce IL-22, which in turn affects keratinocytes pathogenically.
Cited by 1 articles
-
Inhibition of Poly(I:C)-Induced Inflammation by Salvianolic Acid A in Skin Keratinocytes
Qing-Ling Zhang, Ri-Hua Jiang, Xue Mei Li, Jung-Woo Ko, Chang Deok Kim, Ming Ji Zhu, Jeung-Hoon Lee
Ann Dermatol. 2019;31(3):279-285. doi: 10.5021/ad.2019.31.3.279.
Reference
-
1. Li Y, Begovich AB. Unraveling the genetics of complex diseases: susceptibility genes for rheumatoid arthritis and psoriasis. Semin Immunol. 2009; 21:318–327.
Article2. Nakajima H, Nakajima K, Tarutani M, Morishige R, Sano S. Kinetics of circulating Th17 cytokines and adipokines in psoriasis patients. Arch Dermatol Res. 2011; 303:451–455.
Article3. Becher B, Pantelyushin S. Hiding under the skin: interleukin-17-producing γδ T cells go under the skin? Nat Med. 2012; 18:1748–1750.
Article4. Pappu R, Rutz S, Ouyang W. Regulation of epithelial immunity by IL-17 family cytokines. Trends Immunol. 2012; 33:343–349.
Article5. Iwakura Y, Nakae S, Saijo S, Ishigame H. The roles of IL-17A in inflammatory immune responses and host defense against pathogens. Immunol Rev. 2008; 226:57–79.
Article6. Truchetet ME, Mossalayi MD, Boniface K. IL-17 in the rheumatologist's line of sight. Biomed Res Int. 2013; DOI: 10.1155/2013/295132.
Article7. Iwakura Y, Ishigame H, Saijo S, Nakae S. Functional specialization of interleukin-17 family members. Immunity. 2011; 34:149–162.
Article8. Zhang L, Yang XQ, Cheng J, Hui RS, Gao TW. Increased Th17 cells are accompanied by FoxP3(+) Treg cell accumulation and correlated with psoriasis disease severity. Clin Immunol. 2010; 135:108–117.
Article9. Rich P, Sigurgeirsson B, Thaci D, Ortonne JP, Paul C, Schopf RE, et al. Secukinumab induction and maintenance therapy in moderate-to-severe plaque psoriasis: a randomized, double-blind, placebo-controlled, phase II regimen-finding study. Br J Dermatol. 2013; 168:402–411.
Article10. Trinchieri G, Sher A. Cooperation of Toll-like receptor signals in innate immune defence. Nat Rev Immunol. 2007; 7:179–190.
Article11. Medzhitov R. Toll-like receptors and innate immunity. Nat Rev Immunol. 2001; 1:135–145.
Article12. Iwasaki A, Medzhitov R. Toll-like receptor control of the adaptive immune responses. Nat Immunol. 2004; 5:987–995.
Article13. McInturff JE, Modlin RL, Kim J. The role of toll-like receptors in the pathogenesis and treatment of dermatological disease. J Invest Dermatol. 2005; 125:1–8.
Article14. Kawai K. Expression of functional toll-like receptors on cultured human epidermal keratinocytes. J Invest Dermatol. 2003; 121:217.
Article15. Miller LS, Sørensen OE, Liu PT, Jalian HR, Eshtiaghpour D, Behmanesh BE, et al. TGF-alpha regulates TLR expression and function on epidermal keratinocytes. J Immunol. 2005; 174:6137–6143.
Article16. Li ZJ, Sohn KC, Choi DK, Shi G, Hong D, Lee HE, et al. Roles of TLR7 in activation of NF-κB signaling of keratinocytes by imiquimod. PLoS One. 2013; 8:e77159.
Article17. Köllisch G, Kalali BN, Voelcker V, Wallich R, Behrendt H, Ring J, et al. Various members of the Toll-like receptor family contribute to the innate immune response of human epidermal keratinocytes. Immunology. 2005; 114:531–541.
Article18. Lebre MC, van der Aar AM, van Baarsen L, van Capel TM, Schuitemaker JH, Kapsenberg ML, et al. Human keratinocytes express functional Toll-like receptor 3, 4, 5, and 9. J Invest Dermatol. 2007; 127:331–341.
Article19. Res PC, Piskin G, de Boer OJ, van der Loos CM, Teeling P, Bos JD, et al. Overrepresentation of IL-17A and IL-22 producing CD8 T cells in lesional skin suggests their involvement in the pathogenesis of psoriasis. PLoS One. 2010; 5:e14108.
Article20. Garber K. Genetics: deep exploration. Nature. 2012; 492:S56–S57.
Article21. Balato A, Lembo S, Mattii M, Schiattarella M, Marino R, De Paulis A, et al. IL-33 is secreted by psoriatic keratinocytes and induces pro-inflammatory cytokines via keratinocyte and mast cell activation. Exp Dermatol. 2012; 21:892–894.
Article22. Gutowska-Owsiak D, Ogg GS. The epidermis as an adjuvant. J Invest Dermatol. 2012; 132:940–948.
Article23. Miller LS, Modlin RL. Toll-like receptors in the skin. Semin Immunopathol. 2007; 29:15–26.
Article24. Terhorst D, Kalali BN, Ollert M, Ring J, Mempel M. The role of toll-like receptors in host defenses and their relevance to dermatologic diseases. Am J Clin Dermatol. 2010; 11:1–10.
Article25. Mogensen TH. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev. 2009; 22:240–273.
Article26. Oeckinghaus A, Hayden MS, Ghosh S. Crosstalk in NF-κB signaling pathways. Nat Immunol. 2011; 12:695–708.
Article27. Jeong MS, Kim JY, Lee HI, Seo SJ. Calcitriol may down-regulate mRNA over-expression of toll-like receptor-2 and -4, LL-37 and proinflammatory cytokines in cultured human keratinocytes. Ann Dermatol. 2014; 26:296–302.
Article28. Venugopal PG, Nutman TB, Semnani RT. Activation and regulation of toll-like receptors (TLRs) by helminth parasites. Immunol Res. 2009; 43:252–263.
Article29. Schauber J, Dombrowski Y, Besch R. Pathogenic DNA: cytosolic DNA promotes inflammation in psoriasis. Cell Cycle. 2011; 10:3038–3039.
Article30. Krueger JG, Bowcock A. Psoriasis pathophysiology: current concepts of pathogenesis. Ann Rheum Dis. 2005; 64:Suppl 2. ii30–ii36.
Article31. Boniface K, Guignouard E, Pedretti N, Garcia M, Delwail A, Bernard FX, et al. A role for T cell-derived interleukin 22 in psoriatic skin inflammation. Clin Exp Immunol. 2007; 150:407–415.
Article32. Dyring-Andersen B, Skov L, Løvendorf MB, Bzorek M, Søndergaard K, Lauritsen JP, et al. CD4(+) T cells producing interleukin (IL)-17, IL-22 and interferon-γ are major effector T cells in nickel allergy. Contact Dermatitis. 2013; 68:339–347.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Effects of Interleukin 4 on the Production of Interleukin 6 in Human Keratinocytes
- I-Kappa-B-Zeta Regulates Interleukin-17A/ Tumor Necrosis Factor-Alpha Mediated Synergistic Induction of Interleukin-19 and Interleukin-20 in Humane Keratinocytes
- Effects of Thyroid Hormone on Preduction of Interleukin-6 and Interleukin-11 in Human Bone Marrow Stromal Cells
- Interleukin-17 and Interleukin-22 Induced Proinflammatory Cytokine Production in Keratinocytes via Inhibitor of Nuclear Factor kappaB Kinase-alpha Expression
- The effect of IL-1 from keratinocytes on production of IL-2 by peripheral lymphocytes
