Inhibition of SKP2 Sensitizes Bromocriptine-Induced Apoptosis in Human Prolactinoma Cells
- Affiliations
-
- 1Department of Neurosurgery, Shanghai Institute of Neurosurgery, Changzheng Hospital, Second Military Medical University, Shanghai, China. dxuehua_changzheng@163.com
- 2Department of Neurosurgery, The 411th Hospital of PLA, Shanghai, China.
- KMID: 2378108
- DOI: http://doi.org/10.4143/crt.2016.017
Abstract
- PURPOSE
Prolactinoma (prolactin-secreting pituitary adenoma) is one of the most common estrogen-related functional pituitary tumors. As an agonist of the dopamine D2 receptor, bromocriptine is used widely to inhibit prolactinoma progression. On the other hand, it is not always effective in clinical application. Although a dopamine D2 receptor deficiency contributes to the impaired efficiency of bromocriptine therapy to some extent, it is unknown whether there some other underlying mechanisms leading to bromocriptine resistance in prolactinoma treatment. That is the main point addressed in this project.
MATERIALS AND METHODS
Human prolactinoma samples were used to analyze the S-phase kinase associated protein 2 (SKP2) expression level. Nutlin-3/adriamycin/cisplatin-treated GH3 and MMQ cells were used to analyze apoptosis in SKP2 overexpression or knockdown cells. SKP2 expression and the interaction partners of SKP2 were also detected after a bromocriptine treatment in 293T. Apoptosis was analyzed in C25 and bromocriptine-treated GH3 cells.
RESULTS
Compared to normal pituitary samples, most prolactinoma samples exhibit higher levels of SKP2 expression, which could inhibit apoptosis in a p53-dependent manner. In addition, the bromocriptine treatment prolonged the half-life of SKP2 and resulted in SKP2 overexpression to a greater extent, which in turn compromised its pro-apoptotic effect. As a result, the bromocriptine treatment combined with C25 (a SKP2 inhibitor) led to the maximal apoptosis of human prolactinoma cells.
CONCLUSION
These findings indicated that SKP2 inhibition sensitized the prolactinoma cells to bromocriptine and helped promote apoptosis. Moreover, a combined treatment of bromocriptine and C25 may contribute to the maximal apoptosis of human prolactinoma cells.
Keyword
MeSH Terms
Figure
-

Fig. 1. SKP2 was up-regulated in pituitary prolactinoma. (A) Real-time RT-PCR analysis for SKP2 expression in normal and different types of adenomatous pituitary tissues as indicated. (B) Representative western blotting showing protein expression of the SKP2 in pituitary adenoma samples. Samples with the two highest SKP2 gene expressions in Fig. 1A of each group were selected for protein analysis. (C) Immunohistochemistry of PRL and normal pituitary adenoma samples to detect the level of SKP2. SKP2, S-phase kinase associated protein 2; RT-PCR, reverse transcription polymerase chain reaction; PRL, prolactin; GH, growth hormone; ACTH, adreno-cortico-tropic-hormone; NFPA, non-functioning pituitary adenoma.

Fig. 2. SKP2 overexpression suppresses apoptosis through p53 in GH3 cells. (A) GH3 cells infected with a lentiviral vector stably expressing the rat SKP2-inducible system. SKP2 was induced by 1 μg/mL doxycycline for 24 hours, and cells were then lysed and collected for immunoblotting analysis. (B, C). Twenty-four hours after inducing SKP2, the cells were treated with 5 μM nutlin-3 for 48 hours, and then lysed and collected either for immunoblotting analysis using the antibodies as indicated (B) or for quantitative real-time PCR analysis (C). (D) Twenty-four hours after inducing SKP2 expression, a reporter containing a synthetic p53-binding site (p53-luc) or reporter containing a mutation on the p53-binding site (p53MUT-luc) was transfected into the GH3 cells, together with a control reporter containing renilla luciferase. Twelve hours after transfection, the cells were treated with different concentrations of nutlin-3, as indicated for 48 hours, and the cells were then lysed and collected for luciferase activity analysis. (E) Twenty-four hours after SKP2 induction, the cells were treated with 10 μM nutlin-3 for 48 hours; apoptosis was determined and is shown by measuring the relative caspase 3/7 activity. SKP2, S-phase kinase associated protein 2; PCR, polymerase chain reaction; Dox, doxycycline. *p < 0.05, **p < 0.01, ***p < 0.001; ns, no significance.
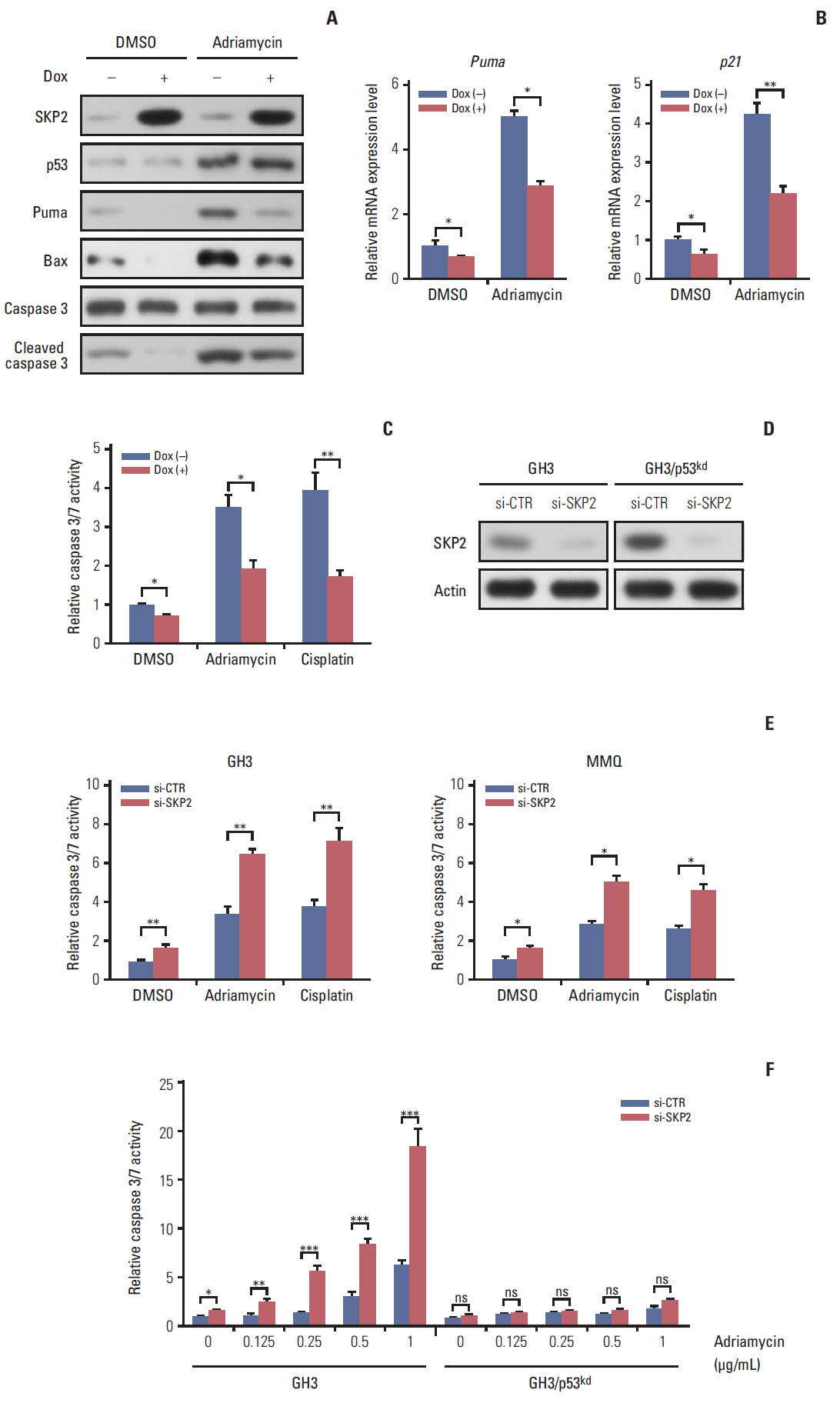
Fig. 3. Skp2 knockdown synergizes with DNA damage induction agents to promote apoptosis in PRL-secreting pituitary cells. (A, B) Twenty-four hours after doxycycline addition, the GH3 cells were treated with 1 μg/mL adriamycin for 48 hours followed by either immunoblotting analysis using the antibodies as indicated (A) or real-time PCR analysis (B). (C) Twenty-four hours after SKP2 induction, the cells were treated with 1 μg/mL adriamycin or 5 μg/mL cisplatin for 48 hours in GH3 cells, followed by relative caspase 3/7 activity determination. (D) A control or Skp2 siRNA was transfected into the wild-type GH3 cells and stable p53 knockdown GH3 cells. Forty-eight hours after transfection, the cells were lysed and collected to analyze SKP2 expression by immunoblotting. (E, F) Twenty-four hours after transfection of the control or Skp2 siRNA, the cells were treated with 1 μg/mL adriamycin or 5 μg/mL cisplatin for 48 hours in the GH3 cells or MMQ cells (E), or indicated adriamycin concentration in wild-type GH3 cells and stable p53 knockdown GH3 cells (F), followed by a determination of the relative caspase 3/7 activity. SKP2, S-phase kinase associated protein 2; PRL, prolactin; PCR, polymerase chain reaction; Dox, doxycycline; DMSO, dimethyl sulfoxide; si-CTR, control siRNA. *p < 0.05, **p < 0.01, ***p < 0.001; ns, no significance.
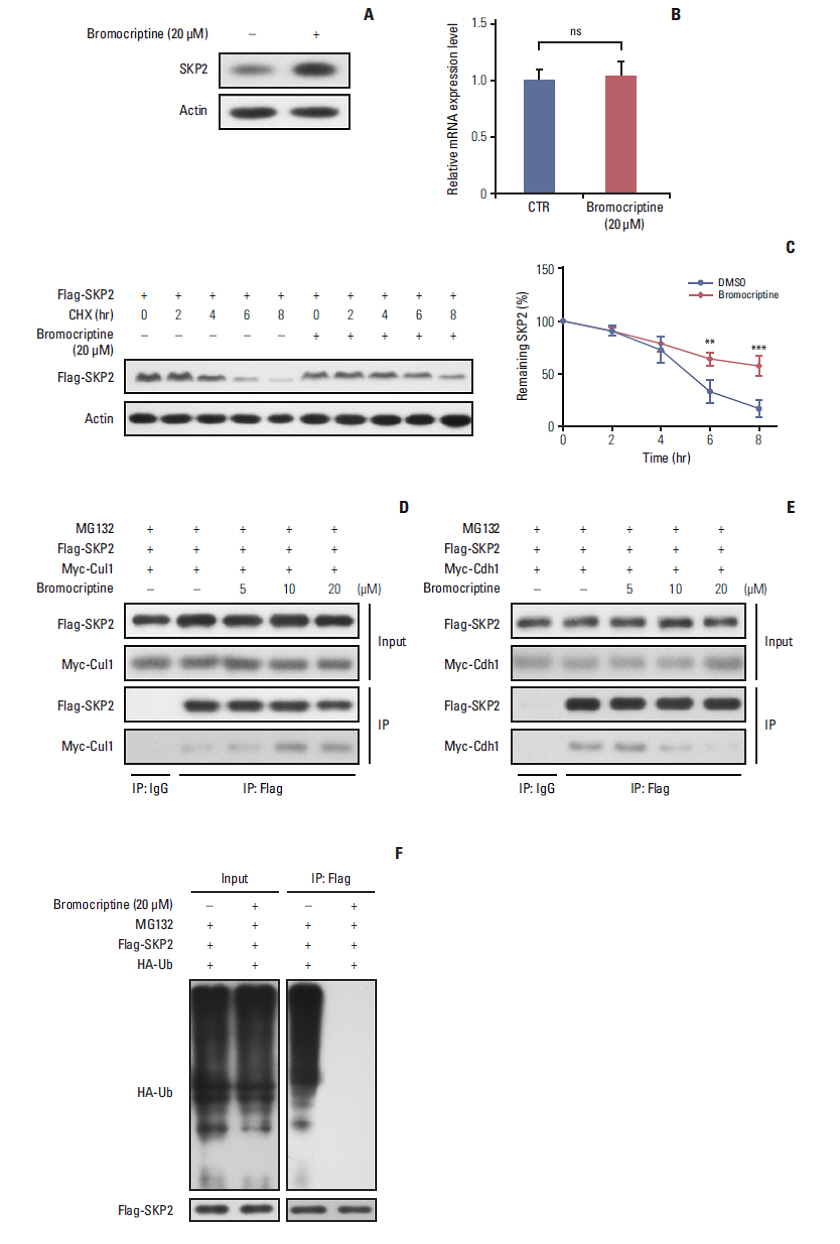
Fig. 4. Bromocriptine stabilizes SKP2 expression by inhibiting SKP2 ubiquitination and degradation. (A, B) SKP2 expression in GH3 cells treated with or without 20 μM bromocriptine for 24 hours, the cells were collected and subjected to immunoblotting (A) or real-time PCR analysis (B). (C) The GH3 cells were transfected with Flag-SKP2. Twenty-four hours after transfection, the cells were treated with 10 μg/mL CHX and 20 μM bromocriptine for the indicated times. The cells were then lysed and subjected to immunoblotting analysis. The SKP2 levels from three independent assays were quantified by densitometry and the measurements were normalized to the start point (0 hour) to calculate the percentage changes following stimulation (C, lower panel). (D, E) the GH3 cells were transfected with Flag-SKP2 and Myc-Cul1 (D) or Flag-SKP2 and Myc-Cdh1 (E). Twenty-four hours after transfection, the cells were treated with 10 μM MG-132 and different concentration of bromocriptine, as indicated for 24 hours, and then cells were subjected to co-immunoprecipitation analysis. (F) The GH3 cells were transfected with Flag-SKP2 and HA-Ub. Twenty-four hours after transfection, the cells were treated with 10 μM MG-132 and 20 μM bromocriptine for 24 hours, and cells were then subjected to co-immunoprecipitation analysis. SKP2, S-phase kinase associated protein 2; CTR, control; PCR, polymerase chain reaction; CHX, cycloheximide; DMSO, dimethyl sulfoxide; HA-Ub, HA-ubiquitin. **p < 0.01, ***p < 0.001; ns, no significance.
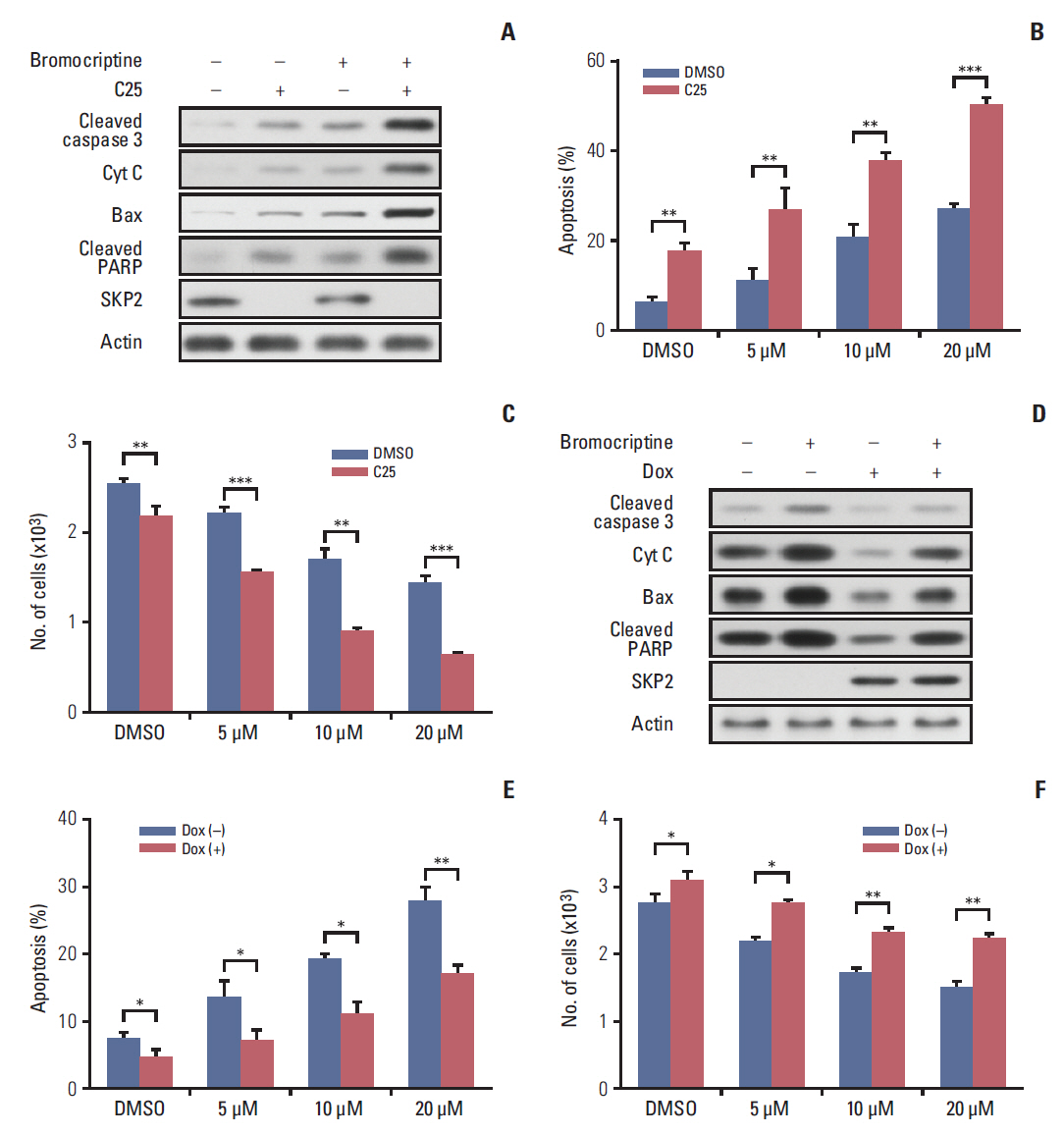
Fig. 5. SKP2 inhibition sensitizes bromocriptine-induced apoptosis in GH3 cells. (A) GH3 cells were treated with 20 μM bromocriptine or 20 μM C25, as indicated for 48 hours. The cells were then lysed and collected for immunoblotting analysis using the indicated antibodies. (B) GH3 cells were treated with 20 μM C25 and different concentrations of bromocriptine, as indicated, for 48 hours. The cells were then lysed and collected for apoptosis determination by annexin-V staining and flow cytometry. (C) The GH3 cells were treated as the same in panel B, and cells were then subjected to determine the number of viable cells. (D) Twenty-four hours after SKP2 induction, the GH3 cells were subjected to 20 μM bromocriptine for 48 hours. The cell lysates were used for immunoblotting analysis using the indicated antibodies. (E) Twenty-four hours after SKP2 induction, the GH3 cells were exposed to the indicated bromocriptine concentrations for a further 48 hours, and cells were then lysed and collected for apoptosis determination by annexin-V staining and flow cytometry. (F) GH3 cells were treated as the same in panel F, and then subjected to a cell viability assay kit to determine the number of viable cells. SKP2, S-phase kinase associated protein 2; Cyt C, cytochrome C; PARP, poly(ADP-ribose) polymerase; DMSO, dimethyl sulfoxide; Dox, doxycycline. *p < 0.05, **p < 0.01, ***p < 0.001
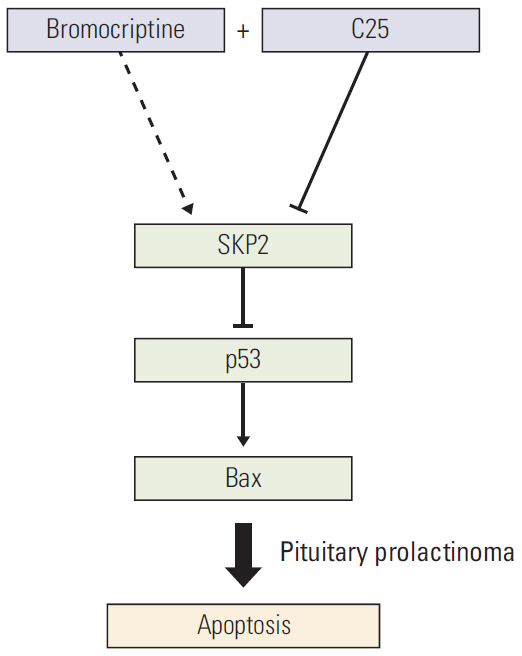
Fig. 6. Schematic model. Schematic model showing how SKP2 overexpression compromised the bromocriptine-induced apoptosis by inhibiting the p53 pathway and the inhibition of SKP2 by C25 sensitizes the bromocriptine-induced apoptosis in PRL-secreting pituitary cells. SKP2, S-phase kinase associated protein 2; PRL, prolactin.
Reference
-
References
1. Glezer A, Bronstein MD. Prolactinomas, cabergoline, and pregnancy. Endocrine. 2014; 47:64–9.
Article2. Molitch ME. Prolactin-secreting tumors: what's new? Expert Rev Anticancer Ther. 2006; 6 Suppl 9:S29–35.
Article3. Booth AK, Gutierrez-Hartmann A. Signaling pathways regulating pituitary lactotrope homeostasis and tumorigenesis. Adv Exp Med Biol. 2015; 846:37–59.
Article4. Heaney AP, Fernando M, Melmed S. Functional role of estrogen in pituitary tumor pathogenesis. J Clin Invest. 2002; 109:277–83.
Article5. Caporali S, Imai M, Altucci L, Cancemi M, Caristi S, Cicatiello L, et al. Distinct signaling pathways mediate stimulation of cell cycle progression and prevention of apoptotic cell death by estrogen in rat pituitary tumor PR1 cells. Mol Biol Cell. 2003; 14:5051–9.
Article6. Jaffrain-Rea ML, Petrangeli E, Ortolani F, Fraioli B, Lise A, Esposito V, et al. Cellular receptors for sex steroids in human pituitary adenomas. J Endocrinol. 1996; 151:175–84.
Article7. Colao A, Di Sarno A, Guerra E, De Leo M, Mentone A, Lombardi G. Drug insight: cabergoline and bromocriptine in the treatment of hyperprolactinemia in men and women. Nat Clin Pract Endocrinol Metab. 2006; 2:200–10.
Article8. Van De Weerdt C, Peers B, Belayew A, Martial JA, Muller M. Far upstream sequences regulate the human prolactin promoter transcription. Neuroendocrinology. 2000; 71:124–37.
Article9. Oh MC, Aghi MK. Dopamine agonist-resistant prolactinomas. J Neurosurg. 2011; 114:1369–79.
Article10. Gillam MP, Molitch ME, Lombardi G, Colao A. Advances in the treatment of prolactinomas. Endocr Rev. 2006; 27:485–534.
Article11. Missale C, Losa M, Boroni F, Giovanelli M, Balsari A, Spano PF. Nerve growth factor and bromocriptine: a sequential therapy for human bromocriptine-resistant prolactinomas. Br J Cancer. 1995; 72:1397–9.
Article12. Gao H, Wang F, Lan X, Li C, Feng J, Bai J, et al. Lower PRDM2 expression is associated with dopamine-agonist resistance and tumor recurrence in prolactinomas. BMC Cancer. 2015; 15:272.
Article13. Cai L, Leng ZG, Guo YH, Lin SJ, Wu ZR, Su ZP, et al. Dopamine agonist resistance-related endocan promotes angiogenesis and cells viability of prolactinomas. Endocrine. 2016; 52:641–51.
Article14. Galluzzi L, Morselli E, Kepp O, Tajeddine N, Kroemer G. Targeting p53 to mitochondria for cancer therapy. Cell Cycle. 2008; 7:1949–55.
Article15. Tokino T, Nakamura Y. The role of p53-target genes in human cancer. Crit Rev Oncol Hematol. 2000; 33:1–6.
Article16. Thapar K, Scheithauer BW, Kovacs K, Pernicone PJ, Laws ER Jr. p53 expression in pituitary adenomas and carcinomas: correlation with invasiveness and tumor growth fractions. Neurosurgery. 1996; 38:765–70.
Article17. Sav A, Rotondo F, Syro LV, Scheithauer BW, Kovacs K. Biomarkers of pituitary neoplasms. Anticancer Res. 2012; 32:4639–54.18. Wang Z, Fukushima H, Inuzuka H, Wan L, Liu P, Gao D, et al. Skp2 is a promising therapeutic target in breast cancer. Front Oncol. 2012; 1:18702.
Article19. Frescas D, Pagano M. Deregulated proteolysis by the F-box proteins SKP2 and beta-TrCP: tipping the scales of cancer. Nat Rev Cancer. 2008; 8:438–49.20. Wei W, Ayad NG, Wan Y, Zhang GJ, Kirschner MW, Kaelin WG Jr. Degradation of the SCF component Skp2 in cell-cycle phase G1 by the anaphase-promoting complex. Nature. 2004; 428:194–8.
Article21. Kitagawa M, Lee SH, McCormick F. Skp2 suppresses p53-dependent apoptosis by inhibiting p300. Mol Cell. 2008; 29:217–31.
Article22. Lee SH, McCormick F. Downregulation of Skp2 and p27/Kip1 synergistically induces apoptosis in T98G glioblastoma cells. J Mol Med (Berl). 2005; 83:296–307.
Article23. Amaral JD, Xavier JM, Steer CJ, Rodrigues CM. The role of p53 in apoptosis. Discov Med. 2010; 9:145–52.24. Shalini S, Dorstyn L, Dawar S, Kumar S. Old, new and emerging functions of caspases. Cell Death Differ. 2015; 22:526–39.
Article25. Kanasaki H, Fukunaga K, Takahashi K, Miyazaki K, Miyamoto E. Involvement of p38 mitogen-activated protein kinase activation in bromocriptine-induced apoptosis in rat pituitary GH3 cells. Biol Reprod. 2000; 62:1486–94.26. Deshaies RJ. SCF and Cullin/Ring H2-based ubiquitin ligases. Annu Rev Cell Dev Biol. 1999; 15:435–67.
Article27. Chan CH, Morrow JK, Li CF, Gao Y, Jin G, Moten A, et al. Pharmacological inactivation of Skp2 SCF ubiquitin ligase restricts cancer stem cell traits and cancer progression. Cell. 2013; 154:556–68.
Article28. Chao W, Xuexin Z, Jun S, Ming C, Hua J, Li G, et al. Effects of resveratrol on cell growth and prolactin synthesis in GH3 cells. Exp Ther Med. 2014; 7:923–8.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- A Case of Pituitary Macroadenoma Concurrently Diagnosed in a Patient Undergoing Antipsychotic Treatment
- Effects of Epidermal growth factor (EGF) on the suppression of GH3 cell growth
- Normal Repetitive Pregnancies and Tumor Regression Induced by Low-Dose Bromocriptine in a Patient with Macroprolactinoma
- A Case of Bromocriptine Resistant Hyperprolactinemia Which was Responsive to Pergolide
- Triptolide sensitizes lung cancer cells to TNF-related apoptosis-inducing ligand (TRAIL)-induced apoptosis by inhibition of NF-kappa B activation
