HIF-1alpha Upregulation due to Depletion of the Free Ubiquitin Pool
- Affiliations
-
- 1Department of Biomedical Sciences, Seoul National University College of Medicine, Seoul, Korea. parkjw@snu.ac.kr
- 2Department of Pharmacology, Seoul National University College of Medicine, Seoul, Korea.
- 3Cancer Research Institute, Seoul National University College of Medicine, Seoul, Korea.
- 4Ischemic/Hypoxic Disease Institute, Seoul National University College of Medicine, Seoul, Korea.
- KMID: 2344170
- DOI: http://doi.org/10.3346/jkms.2015.30.10.1388
Abstract
- Hypoxia-inducible factor 1alpha (HIF-1alpha), which transactivates a variety of hypoxia-induced genes, is rapidly degraded under nomoxia through the hydroxylation-ubiquitination-proteasome pathway. In this study, we addressed how HIF-1alpha is stabilized by proteasome inhibitors. The ubiquitin pool was rapidly reduced after proteasome inhibition, followed by the accumulation of non-ubiquitinated HIF-1alpha. The poly-ubiquitination of HIF-1alpha was resumed by restoration of free ubiquitin, which suggests that the HIF-1alpha stabilization under proteasome inhibition is attributed to depletion of the free ubiquitin pool. Ni2+ and Zn2+ also stabilized HIF-1alpha with depletion of the free ubiquitin pool and these effects of metal ions were attenuated by restoration of free ubiquitin. Ni2+ and Zn2+ may disturb the recycling of free ubiquitin, as MG132 does. Based on these results, the state of the ubiquitin pool seems to be another critical factor determining the cellular level of HIF-1alpha.
Keyword
MeSH Terms
-
Cell Hypoxia/physiology
Cell Line, Tumor
HCT116 Cells
HEK293 Cells
Humans
Hypoxia-Inducible Factor 1, alpha Subunit/biosynthesis/*metabolism
Leupeptins/pharmacology
Nickel/chemistry
Proteasome Endopeptidase Complex/*metabolism
Proteasome Inhibitors/*pharmacology
Ubiquitin/*metabolism
Ubiquitination/*physiology
Up-Regulation
Zinc/chemistry
Hypoxia-Inducible Factor 1, alpha Subunit
Leupeptins
Nickel
Proteasome Endopeptidase Complex
Proteasome Inhibitors
Ubiquitin
Zinc
Figure
-

Fig. 1 Non-ubiquitinated HIF-1α is accumulated under normoxia by proteasome inhibitors. (A) HEK293T, HCT116 and SiHa cells were incubated under normoxic or hypoxic conditions for 8 hr in the absence or presence of 10 µM MG132. Proteins were immunoblotted with the indicated antibodies. (B) HEK293T, HCT116 and SiHa cells were treated with 10 µM MG132 for the indicate times and proteins in cell lysates were immunoblotted. (C) HEK293T cells were treated with 20 µM lactacystin and 100 nM Bortezomib, and HIF-1α and free ubiquitin were immunoblotted.
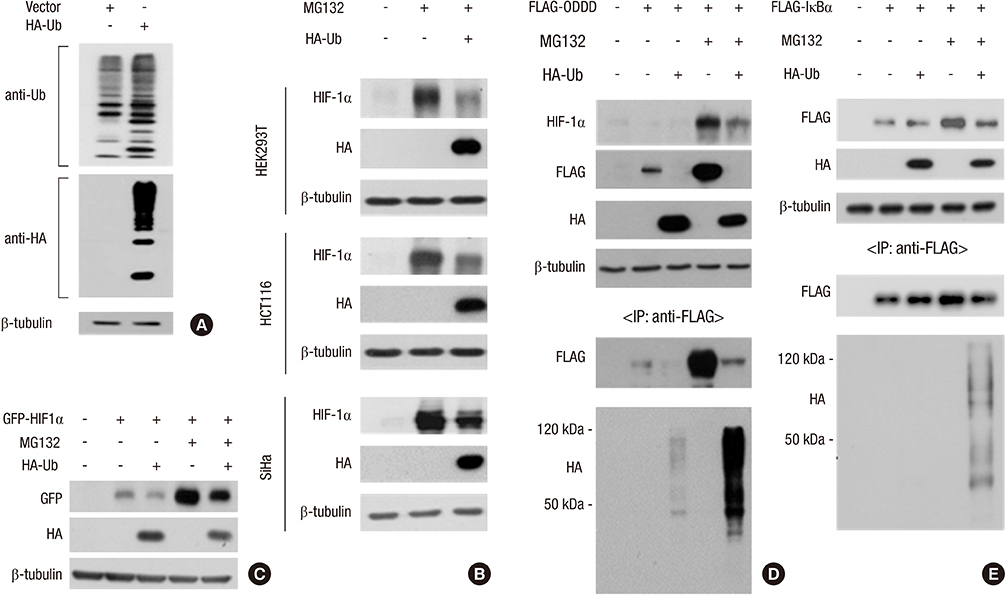
Fig. 2 HIF-1α is ubiquitinated and degraded even under proteasome inhibition by the restoration of free ubiquitin. (A) HEK293T cells were transfected with 1 µg of pcDNA or HA-ubiquitin plasmid. Ubiquitinated proteins in cell lysates were identified using anti-ubiquitin and anti-HA antibodies. (B) HEK293T, HCT116, and SiHa cells were transfected with 1 µg of pcDNA or HA-ubiquitin. After stabilized for 24 hr, cells were treated with 10 µM MG132 for 8 hr, and cell lysates were subjected to immunoblotting. (C) The plasmids for GFP-HIF-1α and HA-ubiquitin were co-transfected into HEK293T cell, and cells were treated with 10 µM MG132 for 8 hr. GFP-HIF-1α and HA-ubiquitin were detected using anti-GFP and anti-HA antibodies, respectively. (D) HEK293T cells were co-transfected with FLAG-ODDD and HA-ubiquitin, and cells were incubated with 10 µM MG132 for 8 hr. The cell lysates were subjected to immunoblotting (upper panel). Proteins in cell lysates were precipitated using anti-FLAG affinity beads and bound proteins were immunoblotted with anti-FLAG or anti-HA antibodies (lower panel). (E) HEK293T cells were co-transfected with FLAG-IκBα and and HA-ubiquitin. After cells were treated with 10 µM MG132 for 8 hr, FLAG-IκBα and free HA-ubiquitin levels were analyzed by immunoblotting (upper panel). Proteins were precipitated using anti-FLAG affinity beads and bound proteins were immunoblotted (lower panel).

Fig. 3 Zinc and nickel ions stabilize HIF-1α by depleting free ubiquitin pool. (A) HEK293T cells were incubated with 100 µM CoCl2, 300 µM ZnCl2, 500 µM NiCl2, or under hypoxic conditions for 1 or 4 hr. HIF-1α and free ubiquitin were immunoblotted. (B) HEK293T cells were transfected with 1 µg of pcDNA or HA-ubiquitin plasmid. After stabilized for 24 hr, cells were incubated with 100 µM CoCl2, 300 µM ZnCl2, 500 µM NiCl2, or under hypoxic conditions for 4 hr. HIF-1α and free ubiquitin in cell lysates were immunoblotted.
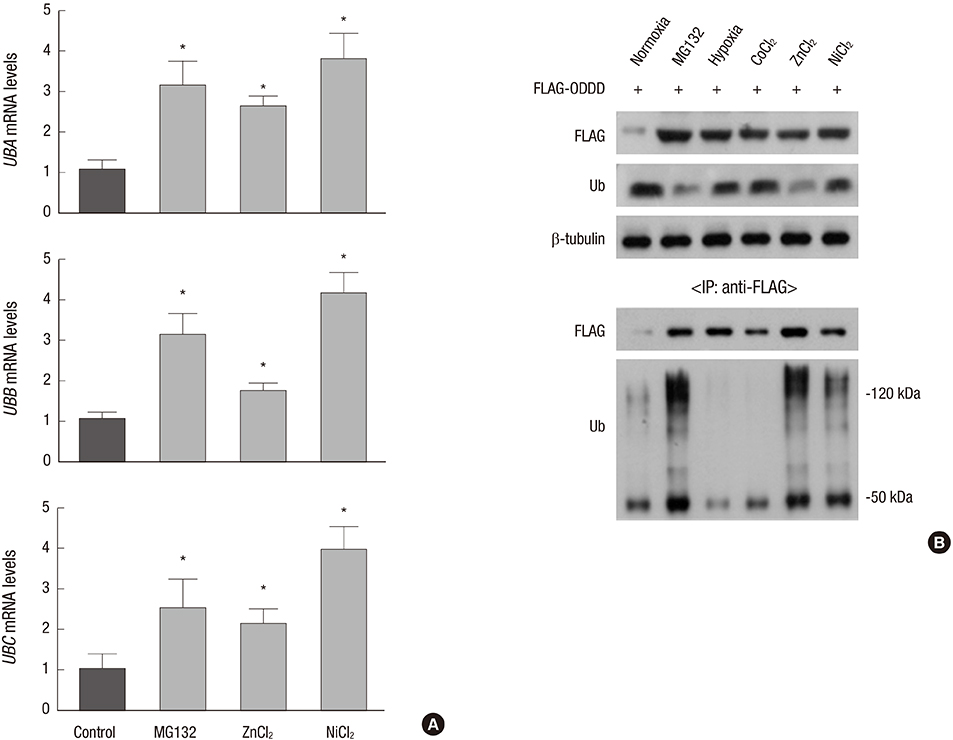
Fig. 4 Zinc and nickel ions induce the accumulation of poly-ubiquitinated peptides. (A) HEK293T cells were treated with 10 µM MG132, 300 µM ZnCl2, or 500 µM NiCl2 for 4 hr. Total mRNAs extracted from the cells were reverse-transcribed and the indicated mRNA levels were quantified using real-time PCR with specific primers for ubiquitin isoforms UBA, UBB, and UBC. Each assay was done in triplication and the result was normalized by the GAPDH level. Final results (means + S.D.) were obtained from five independent experiments. *Denotes P < 0.05 versus the control value. (B) HEK293T cells were transfected with 1 µg of FLAG-ODDD plasmid. After stabilized for 24 hr, cells were incubated with 10 µM MG132, 100 µM CoCl2, 300 µM ZnCl2, 500 µM NiCl2, or under hypoxic conditions for 4 hr. FLAG-ODDD and free ubiquitin in cell lysates were immunoblotted (upper panel). Proteins in cell lysates were precipitated using anti-FLAG affinity beads and bound proteins were immunoblotted (lower panel).
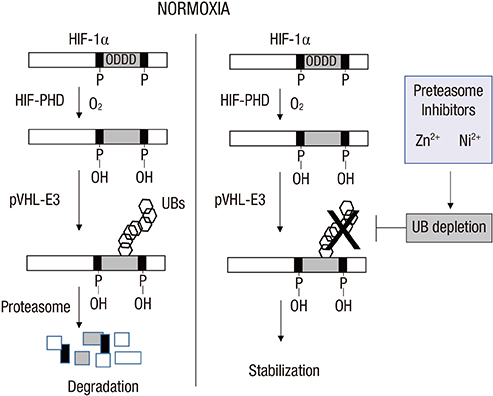
Fig. 5 Hypothesis on HIF-1α stabilization due to depletion of the free ubiquitin pool. Under normoxic conditions, HIF-1α is hydroxylated at it lysine residues through HIF-PHD enzymes and this modification recruits the pVHL/E3 ligase complex to be poly-ubiquitinated, which leads to HIF-1α degradation through the 26S proteasome. Proteasome inhibitors and some metal ions deplete the free ubiquitin pool by disturbing the recycling of free ubiquitin, and then HIF-1α in a non-ubiquitinated form is accumulated even under normoxic conditions. HIF-PHD, HIF-prolyl hydroxylase, VHL, von Hippel Lindau, UB, ubiquitin.
Reference
-
1. Weissman AM. Themes and variations on ubiquitylation. Nat Rev Mol Cell Biol. 2001; 2:169–178.2. Groothuis TA, Dantuma NP, Neefjes J, Salomons FA. Ubiquitin crosstalk connecting cellular processes. Cell Div. 2006; 1:21.3. Thrower JS, Hoffman L, Rechsteiner M, Pickart CM. Recognition of the polyubiquitin proteolytic signal. EMBO J. 2000; 19:94–102.4. Park CW, Ryu KY. Cellular ubiquitin pool dynamics and homeostasis. BMB Rep. 2014; 47:475–482.5. Komander D, Clague MJ, Urbe S. Breaking the chains: structure and function of the deubiquitinases. Nat Rev Mol Cell Biol. 2009; 10:550–563.6. Brahimi-Horn C, Pouyssegur J. When hypoxia signalling meets the ubiquitin-proteasomal pathway, new targets for cancer therapy. Crit Rev Oncol Hematol. 2005; 53:115–123.7. Chun YS, Kim MS, Park JW. Oxygen-dependent and -independent regulation of HIF-1alpha. J Korean Med Sci. 2002; 17:581–588.8. Schofield CJ, Ratcliffe PJ. Oxygen sensing by HIF hydroxylases. Nat Rev Mol Cell Biol. 2004; 5:343–354.9. Kallio PJ, Wilson WJ, O'Brien S, Makino Y, Poellinger L. Regulation of the hypoxia-inducible transcription factor 1alpha by the ubiquitin-proteasome pathway. J Biol Chem. 1999; 274:6519–6525.10. Shin DH, Li SH, Chun YS, Huang LE, Kim MS, Park JW. CITED2 mediates the paradoxical responses of HIF-1alpha to proteasome inhibition. Oncogene. 2008; 27:1939–1944.11. Chun YS, Choi E, Kim GT, Lee MJ, Lee MJ, Lee SE, Kim MS, Park JW. Zinc induces the accumulation of hypoxia-inducible factor (HIF)-1alpha, but inhibits the nuclear translocation of HIF-1beta, causing HIF-1 inactivation. Biochem Biophys Res Commun. 2000; 268:652–656.12. Wang GC, Hazra TK, Mitra S, Lee HM, Englander EW. Mitochondrial DNA damage and a hypoxic response are induced by CoCI2 in rat neuronal PC12 cells. Nucleic Acids Res. 2000; 28:2135–2140.13. Salnikow K, An WG, Melillo G, Blagosklonny MV, Costa M. Nickel-induced transformation shifts the balance between HIF-1 and p53 transcription factors. Carcinogenesis. 1999; 20:1819–1823.14. Xu Q, Farah M, Webster JM, Wojcikiewicz RJ. Bortezomib rapidly suppresses ubiquitin thiolesterification to ubiquitin-conjugating enzymes and inhibits ubiquitination of histones and type I inositol 1,4,5-trisphosphate receptor. Mol Cancer Ther. 2004; 3:1263–1269.15. Melikova MS, Kondratov KA, Kornilova ES. Two different stages of epidermal growth factor (EGF) receptor endocytosis are sensitive to free ubiquitin depletion produced by proteasome inhibitor MG132. Cell Biol Int. 2006; 30:31–43.16. Kim I, Kim CH, Kim JH, Lee J, Choi JJ, Chen ZA, Lee MG, Chung KC, Hsu CY, Ahn YS. Pyrrolidine dithiocarbamate and zinc inhibit proteasome-dependent proteolysis. Exp Cell Res. 2004; 298:229–238.17. Qiu M, Chen Y, Chu Y, Song S, Yang N, Gao J, Wu Z. Zinc ionophores pyrithione inhibits herpes simplex virus replication through interfering with proteasome function and NF-kappaB activation. Antiviral Res. 2013; 100:44–53.18. Cvek B, Milacic V, Taraba J, Dou QP. Ni(II), Cu(II), and Zn(II) diethyldithiocarbamate complexes show various activities against the proteasome in breast cancer cells. J Med Chem. 2008; 51:6256–6258.19. Frezza M, Hindo SS, Tomco D, Allard MM, Cui QC, Heeg MJ, Chen D, Dou QP, Verani CN. Comparative activities of nickel(II) and zinc(II) complexes of asymmetric [NN'O] ligands as 26S proteasome inhibitors. Inorg Chem. 2009; 48:5928–5937.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Phospholipase D2 promotes degradation of hypoxia-inducible factor-1alpha independent of lipase activity
- STAT3 inhibits the degradation of HIF-1alpha by pVHL-mediated ubiquitination
- HIF-1alpha: a Valid Therapeutic Target for Tumor Therapy
- The Relationship between Expression of Hypoxia Inducible Factor-1alpha or Vascular Endothelial Growth Factor and Histopathological Characteristics in Human Transitional Bladder Cancer
- Hypoxia-inducible factor (HIF-1)alpha: its protein stability and biological functions
