The Roles of Reactive Oxygen Species Produced by Contact Allergens and Irritants in Monocyte-derived Dendritic Cells
- Affiliations
-
- 1Department of Dermatology and Cutaneous Biology Research Institute, Yonsei University College of Medicine, Seoul, Korea. mglee@yuhs.ac
- 2Brain Korea 21 Project for Medical Science, Yonsei University College of Medicine, Seoul, Korea.
- 3Department of Dermatology, CHA Bundang Medical Center, CHA University, Seongnam, Korea.
- KMID: 2265359
- DOI: http://doi.org/10.5021/ad.2010.22.3.269
Abstract
- BACKGROUND
Although reactive oxygen species (ROS) have been produced in both mouse bone marrow-derived dendritic cells (DCs) and XS-106 DCs by contact sensitizers and irritants in previous studies, the generation of ROS in human monocyte-derived DCs (MoDCs) and their role in contact hypersensitivity (CHS) has yet to be elucidated.
OBJECTIVE
The purpose of this study was to determine whether contact allergens and irritants induce ROS in MoDCs and, if so, to evaluate the role of contact allergen and irritant induced-ROS in MoDCs in CHS.
METHODS
Production of ROS was measured by 5-(and-6)-chloromethyl-2',7'-dichlorodihydrofluorescein diacetate (CM-H2DCFDA) assay. Surface CD86 and HLA-DR molecules were detected by flow cytometry. Protein carbonylation was detected by Western blotting.
RESULTS
ROS were produced by contact allergens such as dinitrochlorobenzene (DNCB) and thimerosal and the irritant benzalkonium chloride (BKC). DNCB-induced, but not BKC-induced, ROS increased surface CD86 and HLA-DR molecules on MoDCs and induced protein carbonylation. These changes were reduced in the presence of antioxidant N-acetyl cysteine.
CONCLUSION
Our results suggest that DNCB-induced ROS may be different from those induced by irritant BKC. The DNCB-induced ROS may be associated with the CHS response, because they activate surface molecules on DCs that are important for generating immune reactions.
Keyword
MeSH Terms
-
Allergens
Animals
Benzalkonium Compounds
Blotting, Western
Cysteine
Dendritic Cells
Dermatitis, Contact
Dinitrochlorobenzene
Flow Cytometry
HLA-DR Antigens
Humans
Irritants
Mice
Protein Carbonylation
Reactive Oxygen Species
Thimerosal
Allergens
Benzalkonium Compounds
Cysteine
Dinitrochlorobenzene
HLA-DR Antigens
Irritants
Reactive Oxygen Species
Thimerosal
Figure
-
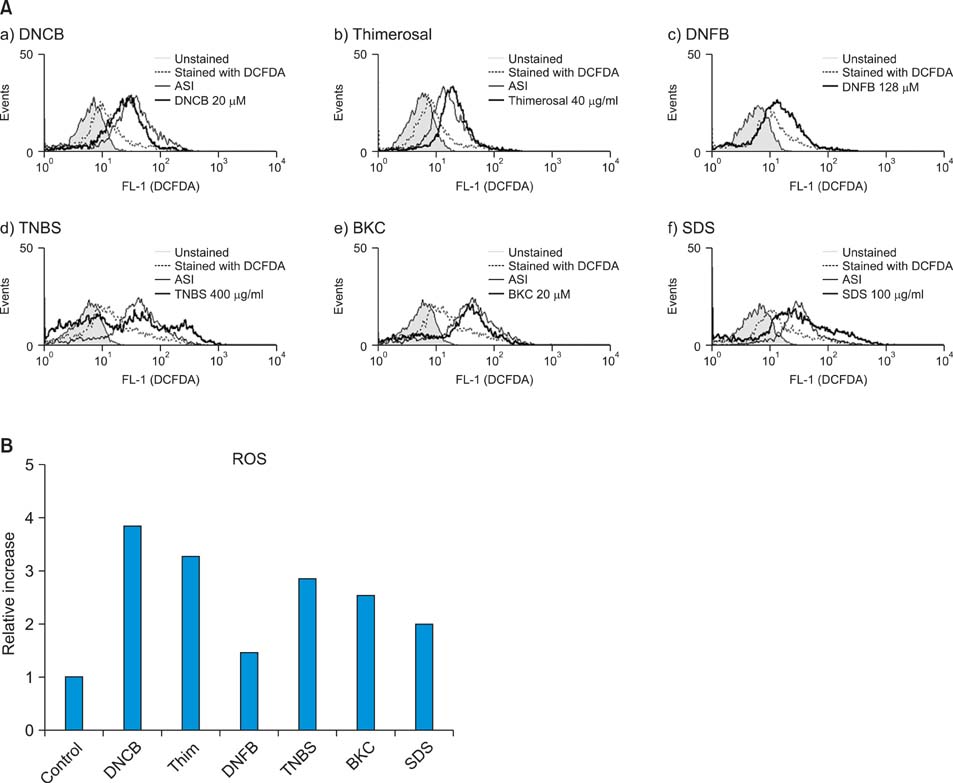
Fig. 1 (A) Monocyte-derived dendritic cells (MoDCs) produce reactive oxygen species (ROS) after incubation with various chemicals including allergens and irritants. Immature MoDCs were incubated for 1 h with sublethal concentration of allergens, such as (a) dinitrochlorobenzene (DNCB), (b) thimerosal (Thim), (c) dinitrofluorobenzene (DNFB) and (d) trinitrobenzene sulfonic acid (TNBS); and irritants, such as (e) benzalkonium chloride (BKC) and (f) sodium dodecyl sulfate (SDS). ROS production was detected by CM-H2DCFDA assay. (A) Filled line represents control cells without CM-H2DCFDA staining; dotted line indicates physiologic ROS level in the cells (DCFDA staining only) without treatment; solid line indicates ATP synthase inhibitor (ASI)-treated cells as a positive control; solid thick line indicates ROS level induced by chemicals. Contact allergens DNCB, TNBS, Thim and DNFB increased ROS production 3.8-fold, 2.9-fold, 3.3-fold, and 1.5-fold, respectively, compared to the control. Irritants BKC and SDS induced ROS significantly (2.6-fold and 2.0-fold, respectively). (B) Relative increase of ROS after incubation with various chemicals including allergens and irritants. The relative increase in ROS was calculated by comparing the mean fluorescence intensity (MFI) of CM-H2DCFDA stained-treatment groups to that of the control group. Of these, the relative increases of ROS production by DNCB, Thim, and DNFB were statistically significant (p-values were 0.031, 0.029, and 0.034, respectively). Of the irritants used, BKC and SDS induced ROS significantly (p-values were 0.028 and 0.013, respectively).
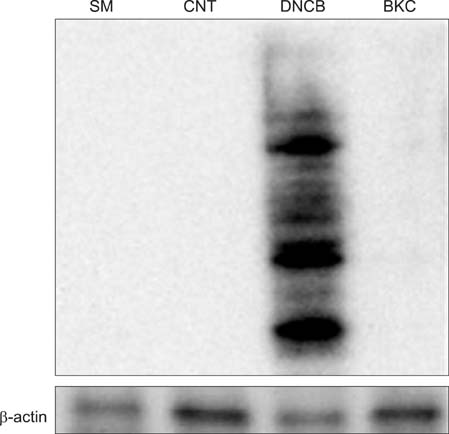
Fig. 2 Contact allergen-DNCB, but not irritant-BKC, induced protein carbonylation. Immature day 6 MoDCs were incubated with 20µm DNCB and 10µM BKC for 6 h. Carbonylated groups were detected by immunoblotting with an anti-DNP antibody. DNCB treated cells but not BKC treated cells showed increased carbonylation compared with the control. Actin is included as a loading control. These data are representative of two independent experiments.
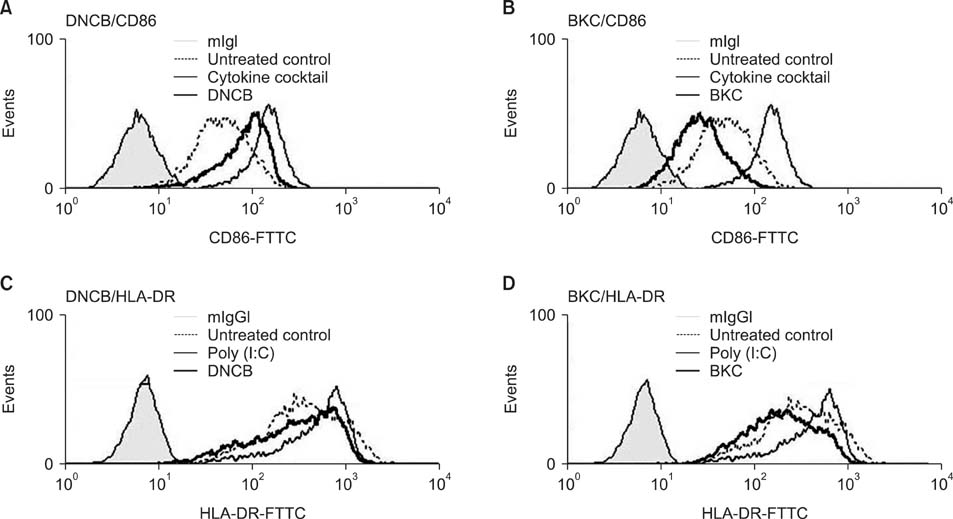
Fig. 3 Augmentation of CD86 and HLA-DR expression on MoDCs by DNCB but not by BKC. MoDCs were treated with 10µM DNCB or 5µM BKC for 24 h and then analyzed by flow cytometry. (A, B) Thick solid line indicates CD86 expression on MoDCs after treatment with DNCB and BKC, respectively; solid narrow line indicates positive control (treatment with cytokine cocktail); filled line indicates untreated DCs; dotted line indicates DCs stained with CD86 only. (C, D) Thick solid line indicates HLA-DR expression on MoDCs after treatment with DNCB and BKC, respectively; solid narrow line indicates positive control, filled line indicates untreated DCs, and dotted line indicates DCs stained with HLA-DR only. CD86 and HLA-DR expression on MoDCs cultured with DNCB was increased 2.5-fold and 1.2-fold respectively; p-values were 0.018 and 0.033, respectively, compared to that of the control.
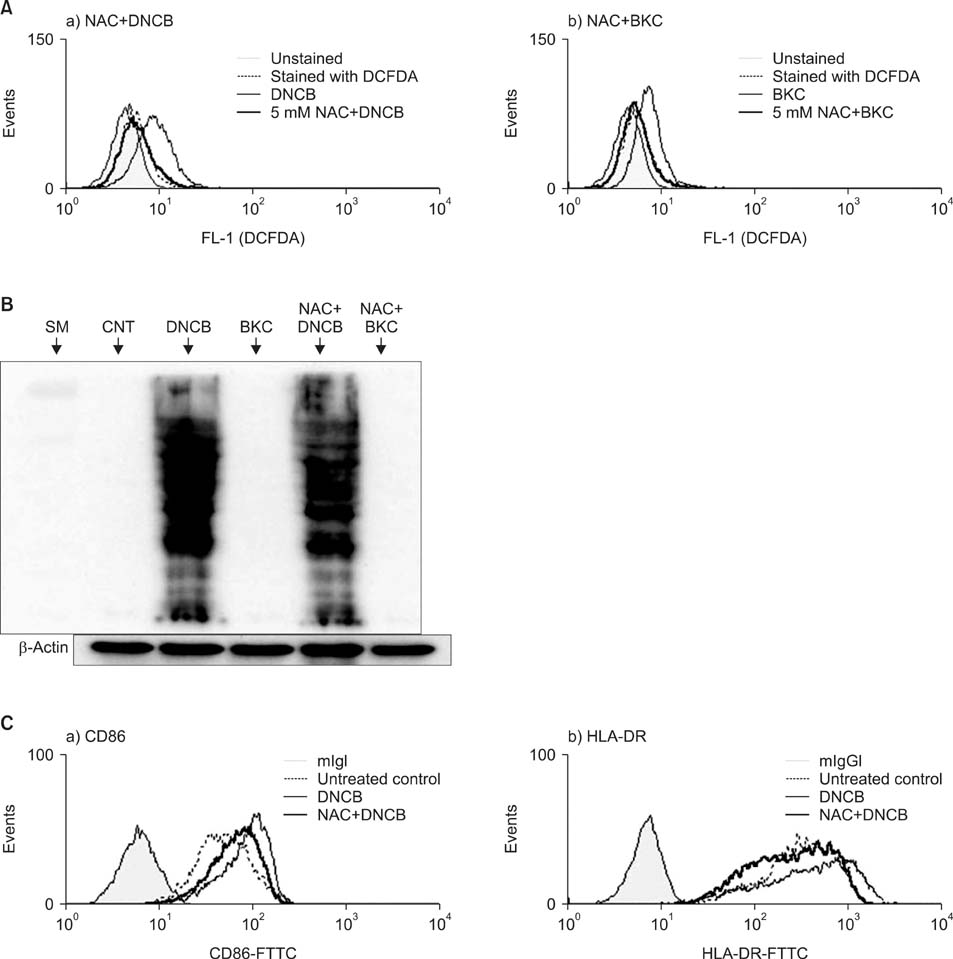
Fig. 4 (A) Hapten and irritant-induced ROS diminished by N-acetyl L-cystein. MoDCs were pre-treated with 5 mM NAC for 30 min and then treated with 30µM DNCB or 20µM BKC. After 1 h they were stained with 3 mM CM-H2DCFDA and analyzed by flow cytometry. The histogram of chemical-treated MoDCs in the presence or the absence of NAC is expressed as a thick solid line or solid line, whereas a filled line or a dotted line indicates histograms of untreated DCs or of DCs stained with CM-H2DCFDA, respectively. Pre-treatment of MoDCs with 5 mM of NAC reduced the ROS production induced by DNCB significantly (the relative ROS increase with DNCB treatment was 3.8-fold while that with pre-treatment with NAC plus DNCB treatment was 0.56-fold. p≤0.0001). BKC-induced ROS production was decreased by pre-treatment with NAC (the relative ROS increase with BKC treatment was 2.6-fold while that with pre-treatment with NAC plus BKC treatment was 0.70-fold). Each experiment was repeated three times and one representative result is shown. (B) These carbonylated proteins were decreased slightly by NAC. Immature day 6 MoDCs were incubated with 20µM DNCB or 10µM BKC in the presence or absence of 5 mM NAC for 30 min. After 6 h, equal amounts of whole cell protein extracts were loaded in SDS gel and carbonylated proteins were detected using DNP antibody by Western blotting. Allergen DNCB but not irritant BKC induced protein carbonylation and the resulting carbonylated protein was decreased slightly by NAC. (C) NAC suppressed the augmentation of CD86 and HLA-DR molecules on MoDCs that were induced by contact allergen DNCB. MoDCs were cultured with DNCB in the presence or absence of pre-treatment 5 mM NAC. After 24 h their surface molecules were analyzed by flow cytometry. Thick solid line indicates the histogram of chemical-treated MoDCs with NAC; narrow solid line indicates treatment without NAC; filled line indicates non-treatment; dotted line indicates treatment with CD86 antibody only. Augmentation of CD86 and HLA-DR by DNCB was decreased by adding 10 mM NAC.
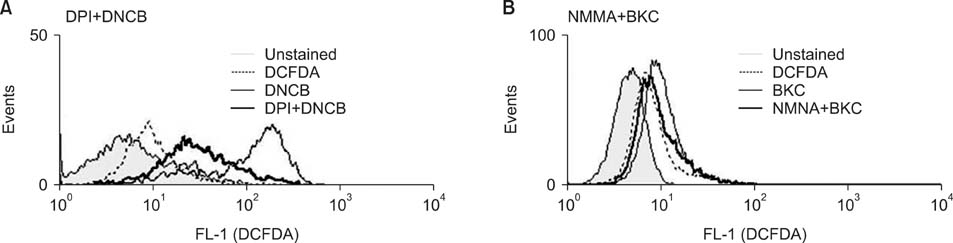
Fig. 5 Inhibition of ROS generating pathway. (A) DNCB induced-ROS blocked by NADPH synthase inhibitor DPI. MoDCs were treated with 20µM DNCB for 1 h in the presence (thick solid line) or absence (solid narrow line) of DPI (a specific blocker of NADPH oxidase). Filled line indicates no staining, while dotted line indicates staining with CM-H2DCFDA only. (B) BKC induced-ROS blocked slightly by iNOS inhibitor NMMA. MoDC treated with 10µM BKC in the absence (solid narrow line) or presence (thick solid line) of NMMA.
Reference
-
1. Rada B, Leto TL. Oxidative innate immune defenses by Nox/Duox family NADPH oxidases. Contrib Microbiol. 2008. 15:164–187.
Article2. Zabucchi G, Bellavite P, Berton G, Dri P. Free radicals generation by the inflammatory cells. Agents Actions Suppl. 1980. 7:159–166.
Article3. Stadtman ER. Protein oxidation and aging. Free Radic Res. 2006. 40:1250–1258.
Article4. Finkel T, Holbrook NJ. Oxidants, oxidative stress and the biology of ageing. Nature. 2000. 408:239–247.
Article5. Waris G, Ahsan H. Reactive oxygen species: role in the development of cancer and various chronic conditions. J Carcinog. 2006. 5:14.
Article6. Valko M, Leibfritz D, Moncol J, Cronin MT, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol. 2007. 39:44–84.
Article7. Bickers DR, Athar M. Oxidative stress in the pathogenesis of skin disease. J Invest Dermatol. 2006. 126:2565–2575.
Article8. Bartosz G. Reactive oxygen species: destroyers or messengers? Biochem Pharmacol. 2009. 77:1303–1315.
Article9. Je JH, Lee TH, Kim DH, Cho YH, Lee JH, Kim SC, et al. Mitochondrial ATP synthase is a target for TNBS-induced protein carbonylation in XS-106 dendritic cells. Proteomics. 2008. 8:2384–2393.
Article10. Nordberg J, Arner ES. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic Biol Med. 2001. 31:1287–1312.
Article11. Fyhrquist-Vanni N, Alenius H, Lauerma A. Contact dermatitis. Dermatol Clin. 2007. 25:613–623.
Article12. Banchereau J, Steinmann RM. Dendritic cells and the control of immunity. Nature. 1998. 392:245–252.
Article13. Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, et al. Immunobiology of dendritic cells. Annu Rev Immunol. 2000. 18:767–811.
Article14. Fukunaga A, Khaskhely NM, Sreevidya CS, Byrne SN, Ullrich SE. Dermal dendritic cells, and not Langerhans cells, play an essential role in inducing an immune response. J Immunol. 2008. 180:3057–3064.
Article15. Grassi F, Dezutter-Dambuyant C, McIlroy D, Jacquet C, Yoneda K, Imamura S, et al. Monocyte-derived dendritic cells have a phenotype comparable to that of dermal dendritic cells and display ultrastructural granules distinct from Birbeck granules. J Leukoc Biol. 1998. 64:484–493.
Article16. Sallusto F, Lanzavecchia A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J Exp Med. 1994. 179:1109–1118.
Article17. Choi GS, Kang JM, Lee MG. Analysis of methods for the generation of dendritic cells from human peripheral blood monocytes. Yonsei Med J. 2000. 41:642–650.
Article18. Kim DS, Kim DH, Byamba D, Lee TH, Cho YH, Lee MG. The production and functions of reactive oxygen species in mouse bone marrow-derived dendritic cells by various haptens and irritants. Korean J Dermatol. 2008. 46:1470–1477.
Article19. Mizuashi M, Ohtani T, Nakagawa S, Aiba S. Redox imbalance induced by contact sensitizers triggers the maturation of dendritic cells. J Invest Dermatol. 2005. 124:579–586.
Article20. Rutault K, Alderman C, Chain BM, Katz DR. Reactive oxygen species activate human peripheral blood dendritic cells. Free Radic Biol Med. 1999. 26:232–238.
Article21. Myhre O, Andersen JM, Aarnes H, Fonnum F. Evaluation of the probes 2',7'-dichlorofluorescin diacetate, luminol, and lucigenin as indicators of reactive species formation. Biochem Pharmacol. 2003. 65:1575–1582.
Article22. Wardman P. Fluorescent and luminescent probes for measurement of oxidative and nitrosative species in cells and tissues: progress, pitfalls, and prospects. Free Radic Biol Med. 2007. 43:995–1022.
Article23. Armstrong JS, Whiteman M. Measurement of reactive oxygen species in cells and mitochondria. Methods Cell Biol. 2007. 80:355–377.
Article24. Yoshida Y, Shimakawa S, Itoh N, Niki E. Action of DCFH and BODIPY as a probe for radical oxidation in hydrophilic and lipophilic domain. Free Radic Res. 2003. 37:861–872.
Article25. Covarrubias L, Hernández-García D, Schnabel D, Salas-Vidal E, Castro-Obregón S. Function of reactive oxygen species during animal development: passive or active? Dev Biol. 2008. 320:1–11.26. Papa L, Gomes E, Rockwell P. Reactive oxygen species induced by proteasome inhibition in neuronal cells mediate mitochondrial dysfunction and a caspase-independent cell death. Apoptosis. 2007. 12:1389–1405.27. Shin MH, Moon YJ, Seo JE, Lee Y, Kim KH, Chung JH. Reactive oxygen species produced by NADPH oxidase, xanthine oxidase, and mitochondrial electron transport system mediate heat shock-induced MMP-1 and MMP-9 expression. Free Radic Biol Med. 2008. 44:635–645.
Article28. Dalle-Donne I, Giustarini D, Colombo R, Rossi R, Milzani A. Protein carbonylation in human diseases. Trends Mol Med. 2003. 9:169–176.
Article29. Arouma OI, Halliwell B, Hoey BM, Butler J. The antioxidant action of N-acetylcysteine: its reaction with hydrogen peroxide, hydroxyl radical, superoxide, and hypochlorous acid. Free Radic Biol Med. 1989. 6:593–597.
Article30. Curtin JF, Donovan M, Cotter TG. Regulation and measurement of oxidative stress in apoptosis. J Immunol Methods. 2002. 265:49–72.
Article31. Stepnik M, Arkusz J. Molecular events associated with dendritic cells activation by contact sensitizers. Int J Occup Med Environ Health. 2003. 16:191–199.
Article32. Aiba S, Manome H, Nakagawa S, Mollah ZU, Mizuashi M, Ohtani T, et al. p38 Mitogen-activated protein kinase and extracellular signal-regulated kinases play distinct roles in the activation of dendritic cells by two representative haptens, NiCl2 and 2,4-dinitrochlorobenzene. J Invest Dermatol. 2003. 120:390–399.
Article33. Trompezinski S, Migdal C, Tailhardat M, Le Varlet B, Courtellemont P, Haftek M, et al. Characterization of early events involved in human dendritic cell maturation induced by sensitizers: cross talk between MAPK signalling pathways. Toxicol Appl Pharmacol. 2008. 230:397–406.
Article34. Watanabe H, Gaide O, Pétrilli V, Martinon F, Contassot E, Roques S, et al. Activation of the IL-1beta-processing inflammasome is involved in contact hypersensitivity. J Invest Dermatol. 2007. 127:1956–1963.
Article35. Matsue H, Edelbaum D, Shalhevet D, Mizumoto N, Yang C, Mummert ME, et al. Generation and function of reactive oxygen species in dendritic cells during antigen presentation. J Immunol. 2003. 171:3010–3018.
Article36. Na K, Kim KE, Park ST, Kim TY. EC-SOD suppresses contact hypersensitivity in mouse skin by impairing Langerhans cell migration. J Invest Dermatol. 2007. 127:1930–1937.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Production and Functions of Reactive Oxygen Species in Mouse Bone Marrow-derived Dendritic Cells by Various Haptens and Irritants
- Effect of Salicylate on the Monocyte Chemoattractant Protein-1 Expression and Intracellular Reactive Oxygen Species Formation in Human Mesangial Cells
- A Case of Epoxy Resin-induced Airborne Allergic Contact Dermatitis
- Keratinocytes-Derived Reactive Oxygen Species Play an Active Role to Induce Type 2 Inflammation of the Skin: A Pathogenic Role of Reactive Oxygen Species at the Early Phase of Atopic Dermatitis
- Do Reactive Oxygen Species Cause Aging?
