Use of Long-term Cultured Embryoid Bodies May Enhance Cardiomyocyte Differentiation by BMP2
- Affiliations
-
- 1Institute of Reproductive Medicine and Population, Medical Research Center, Seoul, Korea. ymchoi@snu.ac.kr
- 2Department of Obstetrics and Gynecology, Seoul National University College of Medicine, Seoul, Korea.
- KMID: 2158201
- DOI: http://doi.org/10.3349/ymj.2008.49.5.819
Abstract
- PURPOSE
Human embryonic stem cells (hESCs) can proliferate for a prolonged period and differentiate into cardiomyocytes in vitro. Recent studies used bone morphogenetic protein 2 (BMP2) to generate cardiomyocytes from hESCs, however, all those studies used early embryoid bodies (EBs) and did not retrieve cardiomyocytes with a high yield. In this study, we treated long-term cultured EBs with BMP2 in order to promote differentiation into cardiomyocytes from hESCs. MATERIALS AND METHODS: hESC lines, including SNUhES3 and SNUhES4, were used in this study. Undifferentiated hESC colonies were detached to form EBs and cultured for up to 30 days. These long-term cultured EBs were differentiated into cardiomyocytes in serum-containing media. In our protocol, BMP2 was applied for 5 days after attachment of EBs. Cardiac specific markers, beating of differentiated cells and electron microscopic (EM) ultrastructures were evaluated and analyzed. RESULTS: Compared to 10-day or 20-day EBs, 30-day EBs showed a higher expression level of cardiac specific markers, Nkx2.5 and a-myosin heavy chain (alphaMHC). Treatment of BMP2 increased expression of cardiac troponin (cTn) I and a-actinin when evaluated at 20 days after attachment of 30-day EBs. Beating of differentiated cells was observed from 7 to 20 days after attachment. Moreover, EM findings demonstrated fine structures such as Z bands in these differentiated cardiomyocytes. These long-term cultured EBs yielded cardiomyocytes with an efficiency of as high as 73.6% when assessed by FACS. CONCLUSION: We demonstrated that the use of long-term cultured EBs may enhance differentiation into cardiomyocytes from hESCs when treated with BMP2.
Keyword
MeSH Terms
Figure
-
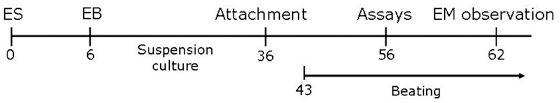
Fig. 1 A schematic presentation of in vitro differentiation strategy for induction of cardiomyocytes from human embryonic stem cells (hESCs), using long-term cultured embryoid bodies (EBs) treated with BMP2.
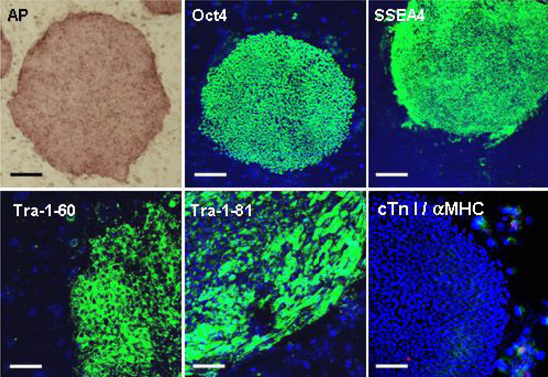
Fig. 2 Expressions of pluripotent hESC- and cardiac specific markers in hESCs. (upper panel) Alkaline phosphatase (AP), Oct4 and SSEA4 were highly expressed in undifferentiated hESCs, Bars: 500 µm. Lower panel) Tra-1-60 and Tra-1-81 were also highly expressed in undifferentiated hESCs. However, cardiac troponin (cTn) I and α-myosin heavy chain (αMHC), cardiac specific markers, were not expressed in undifferentiated hESCs. Merged picture showed only nuclei stained with DAPI. Bars: 200 µm.
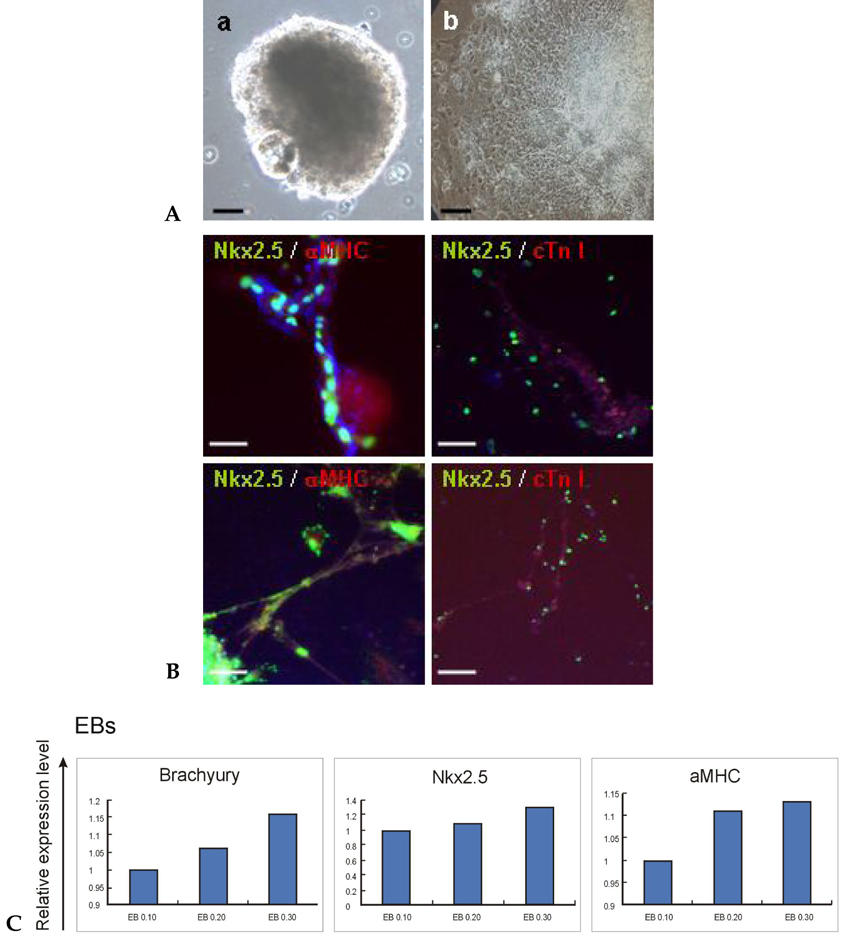
Fig. 3 Evaluation of mesoderm- and cardiac-specific markers in long-term cultured EBs. (A) Morphology of a 30-day EB (a) in suspension and (b) after attachment. Bars: 500 µm. (B) Expressions of cardiac lineage markers in spontaneously differentiated cells from SNUhES3 (upper panel) and SNUhES4 (lower panel). At 20 days after plating, differentiated cells from both hESC lines were positively stained with cardiomyocyte-specific markers, Nkx2.5, αMHC and cTn I, however, their expression was not high and showed a scattered pattern. Bars: 200 µm. (C) Relative expression of mesoderm- and cardiac-lineage markers in long-term cultured EBs from SNUhES3 by Q-PCR. 10-, 20- and 30-day EBs were compared in terms of expression of Brachyury, Nkx2.5 and αMHC, which showed the highest level at day 30 for all three markers.
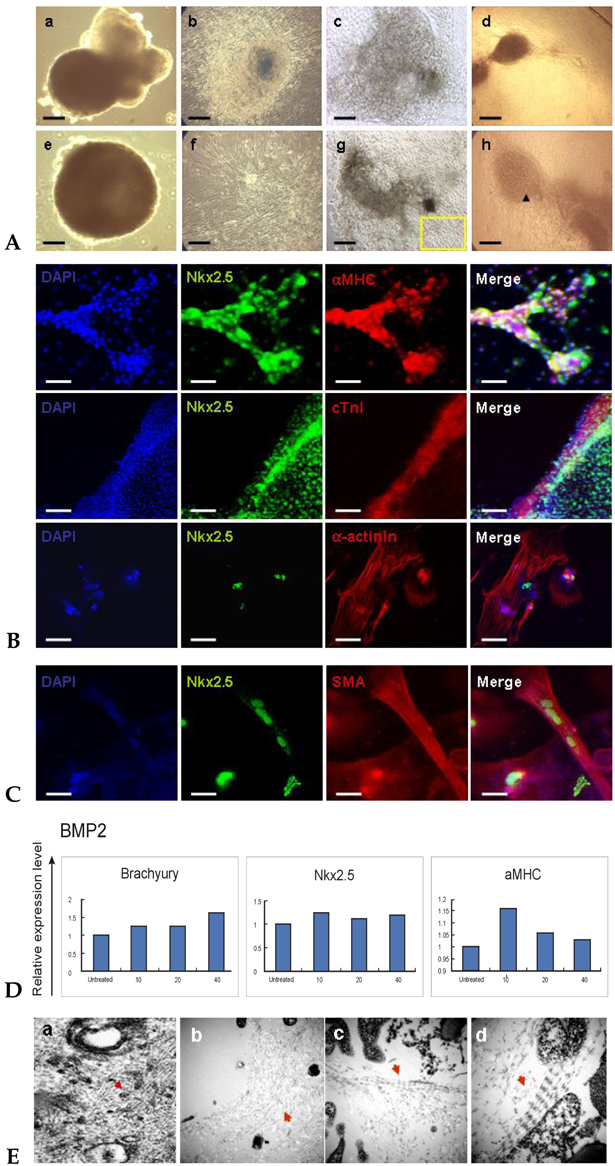
Fig. 4 Evaluation of mesoderm- and cardiac specific markers in cardiomyocytes differentiated from BMP2-treated cells (SNUhES3). (A) Morphologies of long-term cultured embryoid bodies (EBs) and cardiomyocytes differentiated from BMP2-treated cells. Phase contrast microscopic appearance of 30-day EBs (a and e) in suspension and (b and f) after attachment, of hESC-derived cardiomyocytes 4 days (c) and 11 days after attachment (g), and contractile clusters derived from BMP2-treated differentiated cells (d and h) Bar: a-g, 500 µm; (h) 200 µm. (B) Expression of cardiac specific markers in differentiated cardiomyocytes. Differentiated cells were evaluated 20 days after plating and nuclei were stained with DAPI. Top panel, expression of transcription factor Nkx2.5 and protein αMHC; Middle panel, expression of Nkx2.5 and cTn I; Bottom panel, expression of Nkx2.5 and α-actinin (arrow of Fig 4A-h). Bars: 100 µm. (C) Expression of Nkx2.5 and smooth muscle actin (SMA), which is abundant in peripheral cells of cardiomyocyte cluster (yellow box of Fig. 4A-g). Bars: 100 µm. (D) Relative expression of cardiac lineage markers evaluated by Q-PCR. Differentiated cells showed significant expression of Brachyury, Nkx2.5 and αMHC. At 10 ng/mL of BMP2, αMHC expression was the highest, and beating cells were observed in this group (Suppl. Fig. 1). (E) Ultrastructures of differentiated cells observed by transmission electron microscopy (TEM). Red arrows, (a) Z bodies (dense spots), which are the precursors of Z bands. (b) unorganized Z bands. Many unorganized, unconnected bands were observed in many parts. (c and d) organized Z bands. Magnification, a, × 12K; b, × 30K; c, × 60K; d, × 100K.
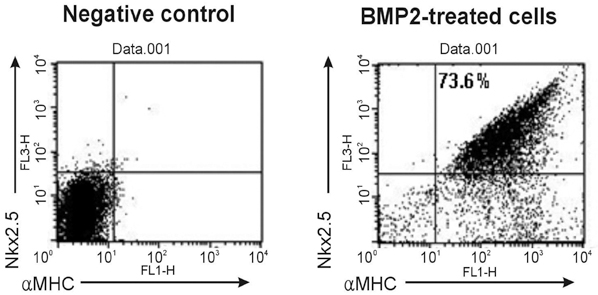
Fig. 5 FACS analysis of cardiac lineage markers, Nkx2.5 and αMHC, in BMP2-induced differentiated cells from plated EBs. As high as 73.6% of differentiated cells were positive for both cardiac markers, evaluated 20 days after plating (Suppl. Fig. 1). Moving image of hESC-derived cardiomyocytes from long-term cultured EBs treated with BMP2. Images were recorded under phase contrast microscope (× 200), and contractile region showed regular contractions with a frequency of 1 beat/second.
Reference
-
1. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, et al. Embryonic stem cell lines derived from human blastocysts. Science. 1998. 282:1145–1147.
Article2. Oh SK, Kim HS, Ahn HJ, Seol HW, Kim YY, Park YB, et al. Derivation and characterization of new human embryonic stem cell lines: SNUhES1, SNUhES2 and SNUhES3. Stem Cells. 2005. 23:211–219.
Article3. Kehat I, Kenyagin-Karsenti D, Snir M, Segev H, Amit M, Gepstein A, et al. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. J Clin Invest. 2001. 108:407–414.
Article4. Xu C, Police S, Rao N, Carpenter MK. Characterization and enrichment of cardiomyocytes derived from human embryonic stem cells. Circ Res. 2002. 91:501–508.
Article5. Mummery C, Ward-van Oostwaard D, Doevendans P, Spijker R, van den Brink S, Hassink R, et al. Differentiation of human embryonic stem cells to cardiomyocytes: role of coculture with visceral endoderm-like cells. Circulation. 2003. 107:2733–2740.
Article6. He JQ, Ma Y, Lee Y, Thomson JA, Kamp TJ. Human embryonic stem cells develop into multiple types of cardiac myocytes: action potential characterization. Circ Res. 2003. 93:32–39.7. Kehat I, Khimovich L, Caspi O, Gepstein A, Shofti R, Arbel G, et al. Electromechanical integration of cardiomyocytes derived from human embryonic stem cells. Nat Biotechnol. 2004. 22:1282–1289.
Article8. Passier R, Oostwaard DW, Snapper J, Kloots J, Hassink R, Kuijk E, et al. Increased cardiomyocyte differentiation from human embryonic stem cells in serum-free cultures. Stem Cells. 2005. 23:772–780.9. Beqqali A, Kloots J, Ward-van Oostwaard D, Mummery C, Passier R. Genome-wide transcriptional profiling of human embryonic stem cells differentiating to cardiomyocytes. Stem Cells. 2006. 24:1956–1967.
Article10. Bettiol E, Sartiani L, Chicha L, Krause KH, Cerbai E, Jaconi ME. Fetal bovine serum enables cardiac differentiation of human embryonic stem cells. Differentiation. 2007. 75:669–681.
Article11. Schlange T, Andrée B, Arnold HH, Brand T. BMP2 is required for early heart development during a distinct time period. Mech Dev. 2000. 91:259–270.
Article12. Yuasa S, Itabashi Y, Koshimizu U, Tanaka T, Sugimura K, Kinoshita M, et al. Transient inhibition of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic stem cells. Nature Biotechnol. 2005. 23:607–611.13. Antin PB, Taylor RG, Yatskievych T. Precardiac mesoderm is specified during gastrulation in quail. Dev Dyn. 1994. 200:144–154.
Article14. Schultheiss TM, Burch JB, Lassar AB. A role for bone morphogenetic proteins in the induction of cardiac myogenesis. Genes Dev. 1997. 11:451–462.
Article15. Tomescot A, Leschik J, Bellamy V, Dubois G, Messas E, Bruneval P, et al. Differentiation in vivo of cardiac committed human embryonic stem cells in postmyocardial infarcted rats. Stem Cells. 2007. 25:2200–2205.
Article16. Pal R, Khanna A. Similar pattern in cardiac differentiation of human embryonic stem cell lines, BG01V and ReiCell® hES1, under low serum concentration supplemented with bone morphogenetic protein-2. Differentiation. 2007. 75:112–122.
Article17. Kwon YD, Oh SK, Kim HS, Ku SY, Kim SH, Choi YM, et al. Cellular manipulation of human embryonic stem cells by TAT-PDX1 protein transduction. Mol Ther. 2005. 12:28–32.
Article18. Oh SK, Kim HS, Park YB, Seol HW, Kim YY, Cho MS, et al. Methods for expansion of human embryonic stem cells. Stem Cells. 2005. 23:605–609.
Article19. Garry DJ, Olson EN. A common progenitor at the heart of development. Cell. 2006. 127:1101–1104.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The effects of BMP4 and BMP2 on Expression of Trophoblast-Specific Genes in Human Embryonic Stem Cells using SNU-hES3 cell line
- Studies on Differentiation of Cardiac Myoblast Induced by Co-culture with Isolated Neonatal Rat Cardiac Myocytes
- Assessment of Developmental Toxicants using Human Embryonic Stem Cells
- SIRT1 Inhibits p53 but not NF-kappaB Transcriptional Activity during Differentiation of Mouse Embryonic Stem Cells into Embryoid Bodies
- Hematopoietic Differentiation of Embryoid Bodies Derived from the Human Embryonic Stem Cell Line SNUhES3 in Co-culture with Human Bone Marrow Stromal Cells
