Priming of Autoreactive CD8+ T Cells Is Inhibited by Immunogenic Peptides Which Are Competitive for Major Histocompatibility Complex Class I Binding
- Affiliations
-
- 1Laboratory of Immunology and Infectious Diseases, Graduate School of Medical Science and Engineering, KAIST, Daejeon 305-701, Korea. ecshin@kaist.ac.kr
- KMID: 2150771
- DOI: http://doi.org/10.4110/in.2013.13.3.86
Abstract
- In the present study, we investigated if priming of autoreactive CD8+ T cells would be inhibited by competitive peptides for major histocompatibility complex (MHC) class I binding. We used a mouse model of vitiligo which is induced by immunization of Kb-binding tyrosinase-related protein 2 (TRP2)-180 peptide. Competitive peptides for Kb binding inhibited IFN-gamma production and proliferation of TRP2-180-specific CD8+ T cells upon ex vivo peptide restimulation, while other MHC class I-binding peptides did not. In mice, the capability of inhibition was influenced by T-cell immunogenicity of the competitive peptides. The competitive peptide with a high T-cell immunogenicity efficiently inhibited priming of TRP2-180-specific CD8+ T cells in vivo, whereas the competitive peptide with a low T-cell immunogenicity did not. Taken together, the inhibition of priming of autoreactive CD8+ T cells depends on not only competition of peptides for MHC class I binding but also competitive peptide-specific CD8+ T cells, suggesting that clonal expansion of autoreactive T cells would be affected by expansion of competitive peptide-specific T cells. This result provides new insights into the development of competitive peptides-based therapy for the treatment of autoimmune diseases.
Keyword
MeSH Terms
Figure
-
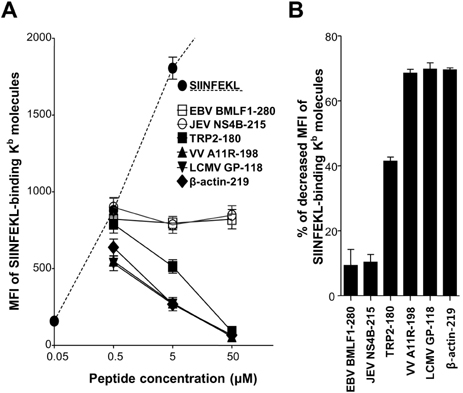
Figure 1 Relative affinity of T cell epitope peptides for Kb binding. Splenocytes of naive C57BL/6 mice were incubated with peptides. Peptide-pulsed splenocytes were fixed with 4% paraformaldehyde, and the density of SIINFEKL-binding Kb molecules on lymphocytes was analyzed by flow cytometry using antibody specific to SIINFEKL-binding Kb molecule (clone 25-D1.16). (A) A dashed line represents the mean fluorescence intensity (MFI) of 25-D1.16-stained lymphocytes pulsed with the indicated concentration of SIINFEKL without additional peptides. Each solid line represents MFI of 25-D1.16-stained lymphocytes pulsed with SIINFEKL at 0.5µM in combination with the indicated concentration of additional peptide. Each dot represents the mean+SEM from 3 independent experiments. (B) The percentage of decreased MFI of SIINFEKL-binding Kb molecules by additional peptides (SIINFEKL at 0.5µM and additional peptide at 5µM) was calculated as follows: [100-MFI of SIINFEKL-binding Kb in SIINFEKL at 0.5µM with additional peptides at 5µM/MFI of SIINFEKL-binding Kb in SIINFEKL at 0.5µM×100]. Each bar graph represents the mean+SEM from 3 independent experiments.
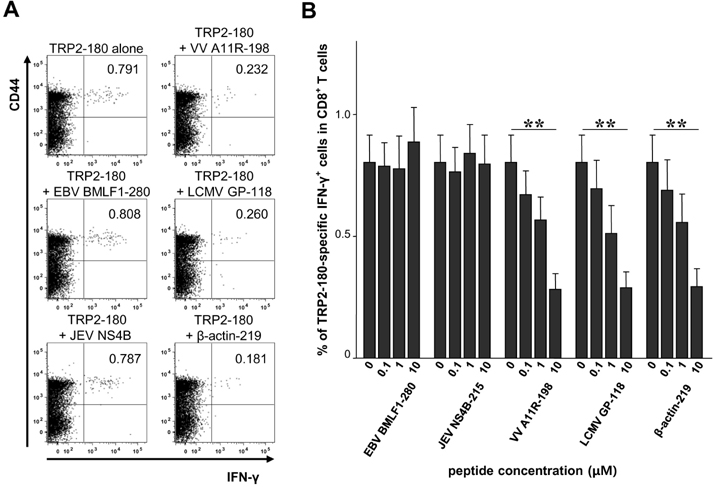
Figure 2 Inhibition of TRP2-180-specific IFN-γ production of CD8+ T cells by competitive peptides for Kb binding in vitro. C57BL/6 mice were immunized with TRP2-180, and splenocytes were harvested and stimulated ex vivo with TRP2-180 peptide alone or in combination with competitive peptides. Then, IFN-γ ICS was performed and analyzed by flow cytometry. (A) The splenocytes were stimulated ex vivo with 0.1µM of TRP2-180 peptide alone or in combination with competitive peptides at 10µM. Representative FACS dot plots for IFN-γ and CD44 are shown in the gate of the CD8+ T cell population. (B) The splenocytes were stimulated ex vivo with TRP2-180 peptide at 0.1µM alone or in combination with the indicated concentration of competitive peptides, and the percentages of TRP2-180-specific IFN-γ+ cells in the CD8+ T-cell population are presented as bar graphs. Each bar graph represents the mean+SEM from 6 independent experiments (**p<0.01).
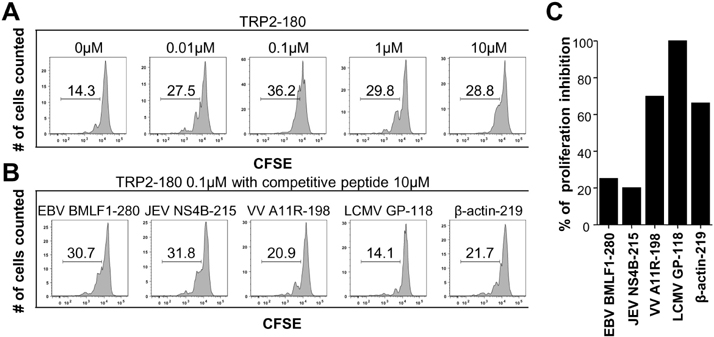
Figure 3 Inhibition of TRP2-180-specific CD8+ T-cell proliferation by competitive peptides for Kb binding in vitro. A TRP2-180-specific T-cell line (CD45.2) labeled with CFSE at 10µM was cocultured with splenocytes (CD45.1) pulsed with TRP2-180 peptide alone or in combination with competitive peptides. Four days after coculture, cocultured cells were stained with anti-CD45.2-APC-H7 and analyzed by flow cytometry. (A) CFSE-labeled T-cell line was cocultured with splenocytes pulsed with the indicated concentration of TRP2-180. The fluorescence intensity of CFSE was analyzed in the gate of the CD45.2+-cell population. (B) CFSE labeled T cell line was cocultured with splenocytes pulsed with TRP2-180 at 0.1µM in combination with additional peptides at 10µM. The fluorescence intensity of CFSE was analyzed in the gate of the CD45.2+-cell population. (C) The percentage of proliferation inhibition by additional peptide was calculated as follows: [100-(% of proliferation in TRP2-180 at 0.1µM with additional peptides-% of proliferation in the absence of TRP2-180)/(% of proliferation in TRP2-180 at 0.1µM-% of proliferation in the absence of TRP2-180)×100], and presented by bar graphs.
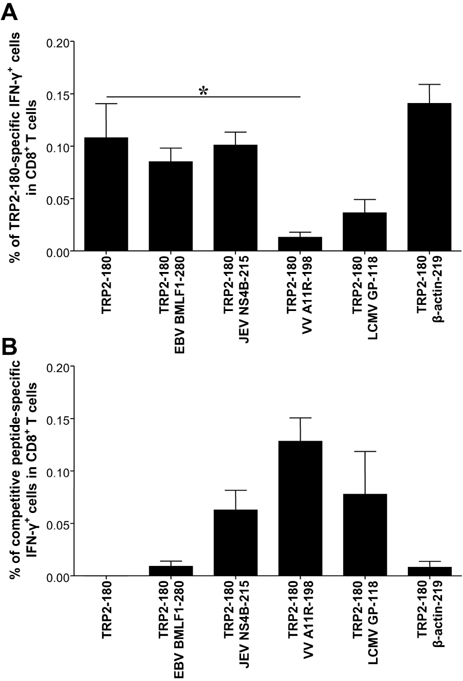
Figure 4 Inhibition of TRP2-180-specific CD8+ T-cell priming by competitive peptides in vivo. C57BL/6 mice were subcutaneously immunized in the footpad with TRP2-180 (5 nmole), LPS, and CpG ODN. One day after the immunization, competitive peptides at 50 nmole were injected into the same footpad without adjuvants twice at 1-day intervals. One week after TRP2-180 immunization, splenocytes were harvested. (A) TRP2-180-specific IFN-γ+ CD8+ T cells were analyzed by ICS and flow cytometry in each group. (B) Competitive peptide-specific IFN-γ+ CD8+ T cells were analyzed by ICS and flow cytometry in each group. Each bar graph represents the mean+SEM from 3 independent experiments with 3 mice per group (*p<0.05).
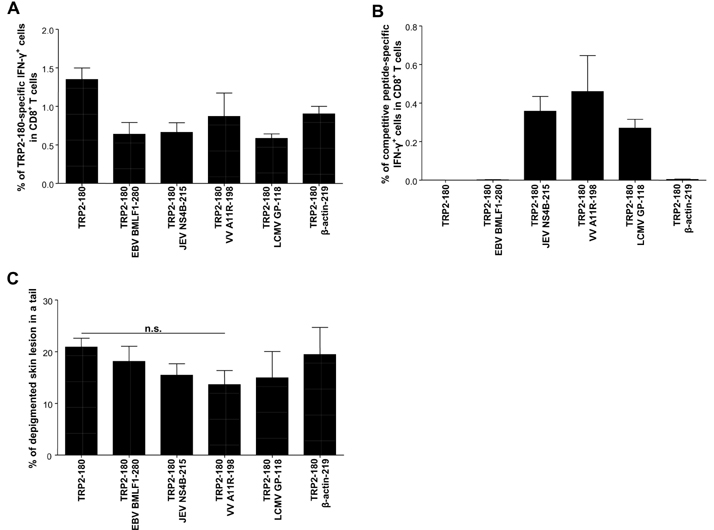
Figure 5 Competitive peptides do not prevent progression of depigmented skin lesions in a mouse model of vitiligo. C57BL/6 mice were subcutaneously immunized in the footpad with TRP2-180 (5 nmole), LPS, and CpG ODN twice at 1-week intervals. One week after the second immunization, TRP2-180 (5 nmole) with the same adjuvants was injected into the tail dermis for the disease induction twice at 1-week intervals. Fifty nmole competitive peptides were injected into the tail dermis 1 day after third and fourth TRP2-180 immunization without adjuvants twice at 1-day intervals. Five weeks after the last TRP2-180 immunization, splenocytes were harvested. (A) TRP2-180-specific IFN-γ+ CD8+ T cells were analyzed by ICS and flow cytometry in each group. (B) Competitive peptide-specific IFN-γ+ CD8+ T cells were analyzed by ICS and flow cytometry in each group. (C) Disease progression was evaluated by the percentage of depigmented skin lesions in the tail in each group. Each bar graph represents the mean+SEM from 3 independent experiments with 3 mice per group (n.s., not significant).
Reference
-
1. Cooper GS, Bynum ML, Somers EC. Recent insights in the epidemiology of autoimmune diseases: improved prevalence estimates and understanding of clustering of diseases. J Autoimmun. 2009; 33:197–207.
Article2. Lee E, Sinha AA. T cell targeted immunotherapy for autoimmune disease. Autoimmunity. 2005; 38:577–596.
Article3. Bjorkman PJ, Saper MA, Samraoui B, Bennett WS, Strominger JL, Wiley DC. Structure of the human class I histocompatibility antigen, HLA-A2. Nature. 1987; 329:506–512.
Article4. Bjorkman PJ, Saper MA, Samraoui B, Bennett WS, Strominger JL, Wiley DC. The foreign antigen binding site and T cell recognition regions of class I histocompatibility antigens. Nature. 1987; 329:512–518.
Article5. Babbitt BP, Matsueda G, Haber E, Unanue ER, Allen PM. Antigenic competition at the level of peptide-Ia binding. Proc Natl Acad Sci U S A. 1986; 83:4509–4513.
Article6. Adorini L, Muller S, Cardinaux F, Lehmann PV, Falcioni F, Nagy ZA. In vivo competition between self peptides and foreign antigens in T-cell activation. Nature. 1988; 334:623–625.
Article7. You S, Cho YH, Byun JS, Shin EC. Melanocyte-specific CD8+ T cells are associated with epidermal depigmentation in a novel mouse model of vitiligo. Clin Exp Immunol. 2013; [Epub ahead of print].
Article8. Porgador A, Yewdell JW, Deng Y, Bennink JR, Germain RN. Localization, quantitation, and in situ detection of specific peptide-MHC class I complexes using a monoclonal antibody. Immunity. 1997; 6:715–726.
Article9. Bloom MB, Perry-Lalley D, Robbins PF, Li Y, el-Gamil M, Rosenberg SA, Yang JC. Identification of tyrosinase-related protein 2 as a tumor rejection antigen for the B16 melanoma. J Exp Med. 1997; 185:453–459.
Article10. Kim SK, Cornberg M, Wang XZ, Chen HD, Selin LK, Welsh RM. Private specificities of CD8 T cell responses control patterns of heterologous immunity. J Exp Med. 2005; 201:523–533.
Article11. Karunakaran KP, Rey-Ladino J, Stoynov N, Berg K, Shen C, Jiang X, Gabel BR, Yu H, Foster LJ, Brunham RC. Immunoproteomic discovery of novel T cell antigens from the obligate intracellular pathogen Chlamydia. J Immunol. 2008; 180:2459–2465.
Article12. Trobaugh DW, Yang L, Ennis FA, Green S. Altered effector functions of virus-specific and virus cross-reactive CD8+ T cells in mice immunized with related flaviviruses. Eur J Immunol. 2010; 40:1315–1327.
Article13. Clute SC, Watkin LB, Cornberg M, Naumov YN, Sullivan JL, Luzuriaga K, Welsh RM, Selin LK. Cross-reactive influenza virus-specific CD8+ T cells contribute to lymphoproliferation in Epstein-Barr virus-associated infectious mononucleosis. J Clin Invest. 2005; 115:3602–3612.
Article14. Yewdell JW, Bennink JR. Immunodominance in major histocompatibility complex class I-restricted T lymphocyte responses. Annu Rev Immunol. 1999; 17:51–88.
Article15. Savir Y, Waysbort N, Antebi YE, Tlusty T, Friedman N. Balancing speed and accuracy of polyclonal T cell activation: a role for extracellular feedback. BMC Syst Biol. 2012; 6:111.
Article16. Curtsinger JM, Lins DC, Mescher MF. CD8+ memory T cells (CD44high, Ly-6C+) are more sensitive than naive cells to (CD44low, Ly-6C-) to TCR/CD8 signaling in response to antigen. J Immunol. 1998; 160:3236–3243.17. Veiga-Fernandes H, Walter U, Bourgeois C, McLean A, Rocha B. Response of naïve and memory CD8+ T cells to antigen stimulation in vivo. Nat Immunol. 2000; 1:47–53.
Article18. Reed C, Katz JM, Hancock K, Balish A, Fry AM. H1N1 Serosurvey Working Group. Prevalence of seropositivity to pandemic influenza A/H1N1 virus in the United States following the 2009 pandemic. PLoS One. 2012; 7:e48187.
Article19. Cannon MJ, Schmid DS, Hyde TB. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev Med Virol. 2010; 20:202–213.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Expression of major histocompatibility complex antigen in Lewis rat cornea
- Relationship between Poor Immunogenicity of HLA-A2-Restricted Peptide Epitopes and Paucity of Naive CD8+ T-Cell Precursors in HLA-A2-Transgenic Mice
- Reconstitution of class I MHC molecules expressed in E. coli and complexed with single antigenic peptides
- Role for CD40 and CD40L Expression in Generating CD8 T Cell Response to Minor Histcompatibility Antigen, H60
- CD43 Expression Regulated by IL-12 Signaling Is Associated with Survival of CD8 T Cells
