CD43 Expression Regulated by IL-12 Signaling Is Associated with Survival of CD8 T Cells
- Affiliations
-
- 1Division of Life and Pharmaceutical Sciences, and Center for Cell Signaling & Drug Discovery Research, Ewha Womans University, Seoul 120-750, Korea. tcell@ewha.ac.kr
- KMID: 1456219
- DOI: http://doi.org/10.4110/in.2010.10.5.153
Abstract
- BACKGROUND
In addition to TCR and costimulatory signals, cytokine signals are required for the differentiation of activated CD8 T cells into memory T cells and their survival. Previously, we have shown that IL-12 priming during initial antigenic stimulation significantly enhanced the survival of activated CD8 T cells and increased the memory cell population. In the present study, we analyzed the mechanisms by which IL-12 priming contributes to activation and survival of CD8 T cells.
METHODS
We observed dramatically decreased expression of CD43 in activated CD8 T cells by IL-12 priming. We purified CD43(lo) and CD43(hi) cells after IL-12 priming and analyzed the function and survival of each population both in vivo and in vitro.
RESULTS
Compared to CD43(hi) effector cells, CD43(lo) effector CD8 T cells exhibited reduced cytolytic activity and lower granzyme B expression but showed increased survival. CD43(lo) effector CD8 T cells also showed increased in vivo expansion after adoptive transfer and antigen challenge. The enhanced survival of CD43(lo) CD8 T cells was also partly associated with CD62L expression.
CONCLUSION
We suggest that CD43 expression regulated by IL-12 priming plays an important role in differentiation and survival of CD8 T cells.
Keyword
Figure
-
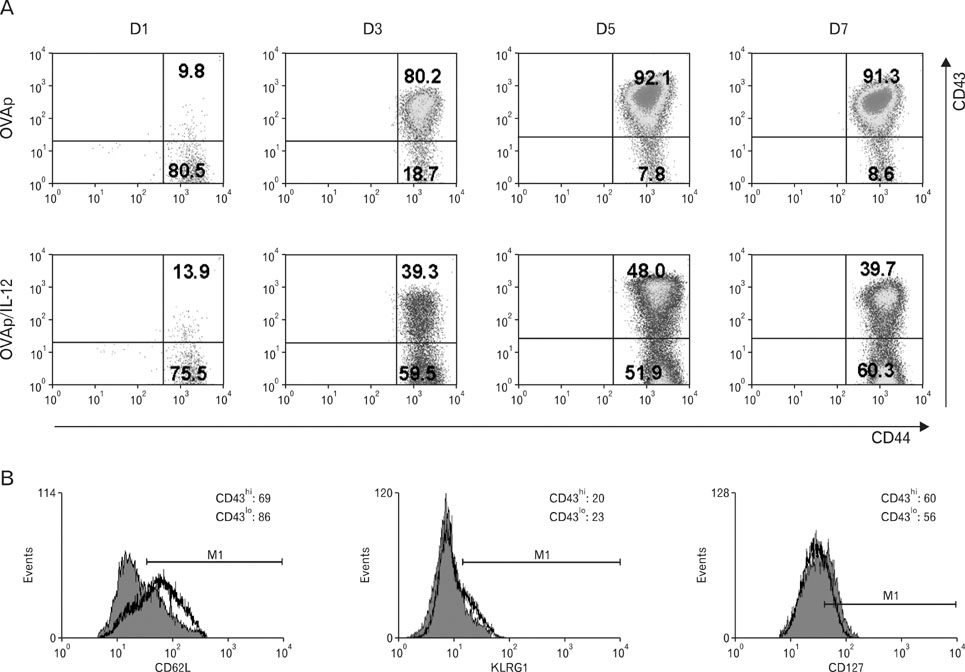
Figure 1 CD43 expression is regulated by initial IL-12 priming during primary CD8 T-cell stimulation. (A) OT-I TCR transgenic cells were stimulated with the OVA peptide (OVAp) in the presence or absence of IL-12. On days 1, 3 and 7, cells were harvested, stained with anti-CD8α, CD44, and CD43, and analyzed by flow cytometry. (B) OT-I cells were stimulated with OVA peptide in the presence of IL-12 (5 ng/ml), and on day 7, cells were harvested, stained with anti-CD8α, CD43 and CD127, CD62L or KLRG1, and analyzed by flow cytometry. Histograms show the expression levels of CD127, CD62L or KLRG1 in the gated CD43hi and CD43lo populations and are shown as the mean±SD. The results shown are representative of three independent experiments with similar results. Filled histogram, CD43hi; open histogram, CD43lo.

Figure 2 CD43lo cells induced by initial IL-12 priming increase their survival rate in vivo, and enhanced survival of CD43lo cells is not due to differential trafficking of transferred cells to various organs. (A) For adoptive transfer experiments, the splenocytes from OT-I TCR transgenic mice were stimulated with the peptide in the presence of IL-12 for 7 days. CD8+CD43hi and CD8+CD43lo cells were MACS-purified and 2×106 cells were i.v. injected into normal C57BL/6 mice. (B) After 3, 10, 17, 34, 45 and 56 days, donor OT-I cells in the blood of recipient mice were identified by CD8α, Kb/OVA-Tet and CD43 staining. (C) On day 7, the frequency of donor OT-I cells in lymphoid and non-lymphoid organs of the recipient mice were measured. The results shown are derived from spleens of 5 mice for each group and are representative of three independent experiments with similar results.
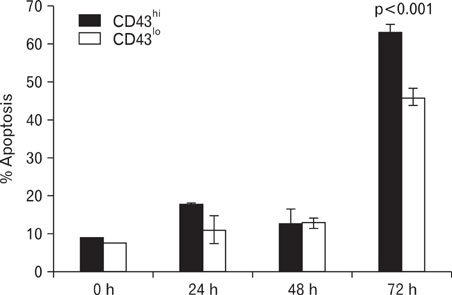
Figure 3 Attenuation of apoptosis in CD43lo population. Spleen cell suspensions from naïve OT-I mice were cultured for 7 days with 100 nM OVAp in the presence of IL-12. Then, activated OT-I cells were MACS-purified and rested for 3 additional days without any stimulant and with IL-2. At the indicated time points, the cells were harvested and apoptotic cell death was determined by Annexin V and 7-AAD double-staining. The samples were assayed in triplicate and the error bars represent the SD values of the mean.
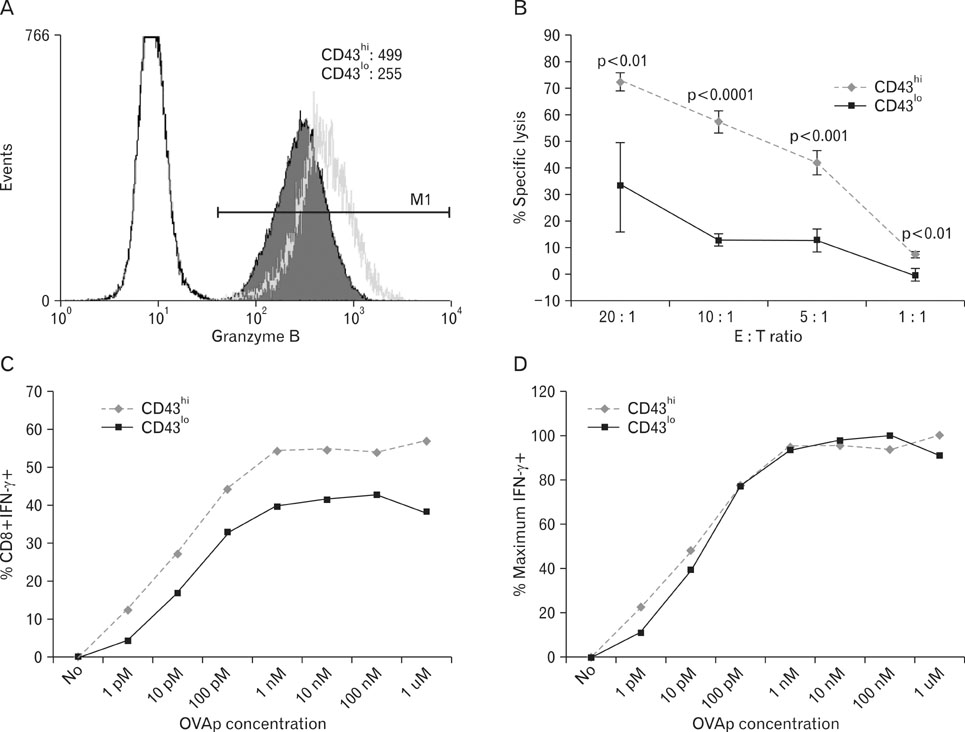
Figure 4 Decreased cytolytic functions in CD43lo population. Spleen cell suspensions from naïve OT-I mice were cultured for 7 days with 100 nM OVAp in the presence of IL-12. Then, activated OT-I cells were MACS-sorted into CD8+CD43hi and CD8+CD43lo populations. (A) The levels of granzyme B in each cell population were determined by intracellular staining. Histograms show the expression of granzyme B in the gated CD43hi and CD43lo subsets and are shown as the mean±SD. The results shown are representative of three independent experiments with similar results. Filled histogram, CD43lo; open green histogram, CD43hi. (B) Peptide-pulsed EL4 target cells (1×104/well) were added to serial dilutions of effector cells (prepared as described above) in 96-well round-bottom plates at E:T ratio of 1:1 to 20:1. After 4 h at 37℃, cytotoxicity was quantified by measurement of cytosolic lactate dehydrogenase (LDH) in the culture supernatant (n=3) using a cytotoxicity detection kit. All experimental procedures and assays were performed two or more times, with similar results. (C) IFN-γ production of purified CD43hi and CD43lo cells was assessed by intracellular IFN-γ staining following 5 h stimulation with 1 pM~1µM OVAp. (D) Data for functional avidity of purified CD43hi and CD43lo cells have been normalized to equate the maximum response obtained after stimulation with the OVAp used to establish the line to 100%.
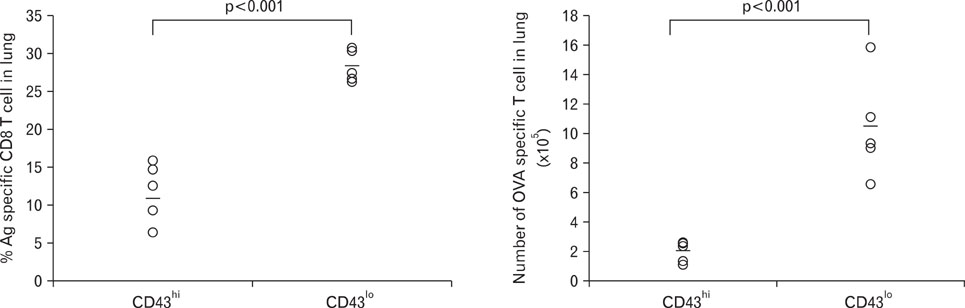
Figure 5 Recall response of CD43lo population is superior to that of CD43hi population. For recall response, the splenocytes from OT-I TCR transgenic mice were stimulated with the antigenic peptide (OVAp) in the presence of IL-12 for 7 days. Then, CD8+CD43hi and CD8+CD43lo cells were separated by MACS. 1×105 purified CD43hi and CD43lo CD8+ T cells in 200µl of PBS were transferred into naïve C57BL/6 mice via tail vein injection and then 1 day after transfer, the mice were intranasally challenged with 2×107 pfu of recombinant adenovirus expressing OVA (rAd/OVA).

Figure 6 Enhanced survival of CD43lo cells is partly associated with CD62L expression. (A) OT-I cells were cultured for 7 days with OVAp in the presence of IL-12. After 7 days, purified CD8+CD62Lhi T cells and CD8+CD62Llo T cells were adoptively transferred into C57BL/6 mice. (B) At the indicated time points, donor cell recovery from the peripheral blood of recipient mice was determined by tetramer staining, and (C) on day 32, the frequency of donor OT-I cells in lymphoid and non-lymphoid organs of the recipient mice was measured.
Cited by 1 articles
-
Baculovirus-based Vaccine Displaying Respiratory Syncytial Virus Glycoprotein Induces Protective Immunity against RSV Infection without Vaccine-Enhanced Disease
Sol Kim, Jun Chang
Immune Netw. 2012;12(1):8-17. doi: 10.4110/in.2012.12.1.8.
Reference
-
1. Kaech SM, Wherry EJ, Ahmed R. Effector and memory T-cell differentiation: implications for vaccine development. Nat Rev Immunol. 2002. 2:251–262.
Article2. Kaech SM, Ahmed R. Memory CD8+ T cell differentiation: initial antigen encounter triggers a developmental program in naïve cells. Nat Immunol. 2001. 2:415–422.
Article3. van Stipdonk MJ, Hardenberg G, Bijker MS, Lemmens EE, Droin NM, Green DR, Schoenberger SP. Dynamic programming of CD8+ T lymphocyte responses. Nat Immunol. 2003. 4:361–365.
Article4. van Stipdonk MJ, Lemmens EE, Schoenberger SP. Naïve CTLs require a single brief period of antigenic stimulation for clonal expansion and differentiation. Nat Immunol. 2001. 2:423–429.
Article5. Wong P, Pamer EG. Cutting edge: antigen-independent CD8 T cell proliferation. J Immunol. 2001. 166:5864–5868.
Article6. Carrio R, Bathe OF, Malek TR. Initial antigen encounter programs CD8+ T cells competent to develop into memory cells that are activated in an antigen-free, IL-7- and IL-15-rich environment. J Immunol. 2004. 172:7315–7323.
Article7. Gett AV, Sallusto F, Lanzavecchia A, Geginat J. T cell fitness determined by signal strength. Nat Immunol. 2003. 4:355–360.
Article8. Langenkamp A, Messi M, Lanzavecchia A, Sallusto F. Kinetics of dendritic cell activation: impact on priming of TH1, TH2 and nonpolarized T cells. Nat Immunol. 2000. 1:311–316.
Article9. Chang J, Cho JH, Lee SW, Choi SY, Ha SJ, Sung YC. IL-12 priming during in vitro antigenic stimulation changes properties of CD8 T cells and increases generation of effector and memory cells. J Immunol. 2004. 172:2818–2826.
Article10. Curtsinger JM, Johnson CM, Mescher MF. CD8 T cell clonal expansion and development of effector function require prolonged exposure to antigen, costimulation, and signal 3 cytokine. J Immunol. 2003. 171:5165–5171.
Article11. Curtsinger JM, Schmidt CS, Mondino A, Lins DC, Kedl RM, Jenkins MK, Mescher MF. Inflammatory cytokines provide a third signal for activation of naive CD4+ and CD8+ T cells. J Immunol. 1999. 162:3256–3262.12. Curtsinger JM, Lins DC, Mescher MF. Signal 3 determines tolerance versus full activation of naive CD8 T cells: dissociating proliferation and development of effector function. J Exp Med. 2003. 197:1141–1151.
Article13. Schluns KS, Williams K, Ma A, Zheng XX, Lefrançois L. Cutting edge: requirement for IL-15 in the generation of primary and memory antigen-specific CD8 T cells. J Immunol. 2002. 168:4827–4831.
Article14. Mescher MF, Curtsinger JM, Agarwal P, Casey KA, Gerner M, Hammerbeck CD, Popescu F, Xiao Z. Signals required for programming effector and memory development by CD8+ T cells. Immunol Rev. 2006. 211:81–92.
Article15. Gahmberg CG, Hayry P, Andersson LC. Characterization of surface glycoproteins of mouse lymphoid cells. J Cell Biol. 1976. 68:642–653.
Article16. Brown WR, Barclay AN, Sunderland CA, Williams AF. Identification of a glycophorin-like molecule at the cell surface of rat thymocytes. Nature. 1981. 289:456–460.
Article17. Moore T, Huang S, Terstappen LW, Bennett M, Kumar V. Expression of CD43 on murine and human pluripotent hematopoietic stem cells. J Immunol. 1994. 153:4978–4987.18. Jones AT, Federsppiel B, Ellies LG, Williams MJ, Burgener R, Duronio V, Smith CA, Takei F, Ziltener HJ. Characterization of the activation-associated isoform of CD43 on murine T lymphocytes. J Immunol. 1994. 153:3426–3439.19. Ellies LG, Jones AT, Williams MJ, Ziltener HJ. Differential regulation of CD43 glycoforms on CD4+ and CD8+ T lymphocytes in graft-versus-host disease. Glycobiology. 1994. 4:885–893.
Article20. Harrington LE, Galvan M, Baum LG, Altman JD, Ahmed R. Differentiating between memory and effector CD8 T cells by altered expression of cell surface O-glycans. J Exp Med. 2000. 191:1241–1246.
Article21. Manjunath N, Correa M, Ardman M, Ardman B. Negative regulation of T-cell adhesion and activation by CD43. Nature. 1995. 377:535–538.
Article22. Stockton BM, Cheng G, Manjunath N, Ardman B, von Andrian UH. Negative regulation of T cell homing by CD43. Immunity. 1998. 8:373–381.
Article23. McEvoy LM, Sun H, Frelinger JG, Butcher EC. Anti-CD43 inhibition of T cell homing. J Exp Med. 1997. 185:1493–1498.
Article24. Ardman B, Sikorski MA, Staunton DE. CD43 interferes with T-lymphocyte adhesion. Proc Natl Acad Sci U S A. 1992. 89:5001–5005.
Article25. Manjunath N, Johnson RS, Staunton DE, Pasqualini R, Ardman B. Targeted disruption of CD43 gene enhances T lymphocyte adhesion. J Immunol. 1993. 151:1528–1534.26. Shaw AS, Dustin ML. Making the T cell receptor go the distance: a topological view of T cell activation. Immunity. 1997. 6:361–369.
Article27. Sperling AI, Sedy JR, Manjunath N, Kupfer A, Ardman B, Burkhardt JK. TCR signaling induces selective exclusion of CD43 from the T cell-antigen-presenting cell contact site. J Immunol. 1998. 161:6459–6462.28. Delon J, Kaibuchi K, Germain RN. Exclusion of CD43 from the immunological synapse is mediated by phosphorylation-regulated relocation of the cytoskeletal adaptor moesin. Immunity. 2001. 15:691–701.
Article29. Roumier A, Olivo-Marin JC, Arpin M, Michel F, Martin M, Mangeat P, Acuto O, Dautry-Varsat A, Alcover A. The membrane-microfilament linker ezrin is involved in the formation of the immunological synapse and in T cell activation. Immunity. 2001. 15:715–728.
Article30. Pedraza-Alva G, Mérida LB, Burakoff SJ, Rosenstein Y. CD43-specific activation of T cells induces association of CD43 to Fyn kinase. J Biol Chem. 1996. 271:27564–27568.
Article31. Santana MA, Pedraza-Alva G, Olivares-Zavaleta N, Madrid-Marina V, Horejsi V, Burakoff SJ, Rosenstein Y. CD43-mediated signals induce DNA binding activity of AP-1, NF-AT, and NFkappa B transcription factors in human T lymphocytes. J Biol Chem. 2000. 275:31460–31468.
Article32. Mattioli I, Dittrich-Breiholz O, Livingstone M, Kracht M, Schmitz ML. Comparative analysis of T-cell costimulation and CD43 activation reveals novel signaling pathways and target genes. Blood. 2004. 104:3302–3304.
Article33. Sperling AI, Green JM, Mosley RL, Smith PL, DiPaolo RJ, Klein JR, Bluestone JA, Thompson CB. CD43 is a murine T cell costimulatory receptor that functions independently of CD28. J Exp Med. 1995. 182:139–146.
Article34. Kyoizumi S, Ohara T, Kusunoki Y, Hayashi T, Koyama K, Tsuyama N. Expression characteristics and stimulatory functions of CD43 in human CD4+ memory T cells: analysis using a monoclonal antibody to CD43 that has a novel lineage specificity. J Immunol. 2004. 172:7246–7253.
Article35. Onami TM, Harrington LE, Williams MA, Galvan M, Larsen CP, Pearson TC, Manjunath N, Baum LG, Pearce BD, Ahmed R. Dynamic regulation of T cell immunity by CD43. J Immunol. 2002. 168:6022–6031.
Article36. Thurman EC, Walker J, Jayaraman S, Manjunath N, Ardman B, Green JM. Regulation of in vitro and in vivo T cell activation by CD43. Int Immunol. 1998. 10:691–701.
Article37. Hikono H, Kohlmeier JE, Takamura S, Wittmer ST, Roberts AD, Woodland DL. Activation phenotype, rather than central-or effector-memory phenotype, predicts the recall efficacy of memory CD8 T cells. J Exp Med. 2007. 204:1625–1636.
Article38. Kaech SM, Tan JT, Wherry EJ, Konieczny BT, Surh CD, Ahmed R. Selective expression of the interleukin 7 receptor identifies effector CD8 T cells that give rise to long-lived memory cells. Nat Immunol. 2003. 4:1191–1198.
Article39. Sarkar S, Kalia V, Haining WN, Konieczny BT, Subramaniam S, Ahmed R. Functional and genomic profiling of effector CD8 T cell subsets with distinct memory fates. J Exp Med. 2008. 205:625–640.
Article40. Hamann D, Baars PA, Rep MH, Hooibrink B, Kerkhof-Garde SR, Klein MR, van Lier RA. Phenotypic and functional separation of memory and effector human CD8 T cells. J Exp Med. 1997. 186:1407–1418.
Article41. Sallusto F, Lenig D, Förster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999. 401:708–712.
Article42. Woodman RC, Johnston B, Hickey MJ, Teoh D, Reinhardt P, Poon BY, Kubes P. The functional paradox of CD43 in leukocyte recruitment: a study using CD43-deficient mice. J Exp Med. 1998. 188:2181–2186.
Article43. Villacres MC, Lacey SF, Auge C, Longmate J, Leedom JM, Diamond DJ. Relevance of peptide avidity to the T cell receptor for cytomegalovirus-specific ex vivo CD8 T cell cytotoxicity. J Infect Dis. 2003. 188:908–918.
Article44. Kalia V, Sarkar S, Gourley TS, Rouse BT, Ahmed R. Differentiation of memory B and T cells. Curr Opin Immunol. 2006. 18:255–264.
Article45. He YW, Bevan MJ. High level expression of CD43 inhibits T cell receptor/CD3-mediated apoptosis. J Exp Med. 1999. 190:1903–1908.
Article46. Allenspach EJ, Cullinan P, Tong J, Tang Q, Tesciuba AG, Cannon JL, Takahashi SM, Morgan R, Burkhardt JK, Sperling AI. ERM-dependent movement of CD43 defines a novel protein complex distal to the immunological synapse. Immunity. 2001. 15:739–750.
Article47. Bagriacik EU, Tang M, Wang HC, Klein JR. CD43 potentiates CD3-induced proliferation of murine intestinal intraepithelial lymphocytes. Immunol Cell Biol. 2001. 79:303–307.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The effect of interleukin 2 on the induction Of Nk 1.1 expression in CD8+ and CD4-CD8-T Cell
- The Roles of CCR7 for the Homing of Memory CD8+ T Cells into Their Survival Niches
- Expression of CD43 in Colorectal Adenocarcinom
- Induction of Unique STAT Heterodimers by IL-21 Provokes IL-1RI Expression on CD8 + T Cells, Resulting in Enhanced IL-1β Dependent Effector Function
- IL-18Ralpha Mediated GATA-3 Induction by Th2 Cells: IL-12 Supports IL-18Ralpha Expression in Th2 Cells
