Cyclooxygenase Inhibitors, Aspirin and Ibuprofen, Inhibit MHC-restricted Antigen Presentation in Dendritic Cells
- Affiliations
-
- 1College of Pharmacy, Chungbuk National University, Cheongju 361-763, Korea. cklee@chungbuk.ac.kr
- 2College of Pharmacy, SahmYook University, Seoul 139-742, Korea.
- KMID: 2150665
- DOI: http://doi.org/10.4110/in.2010.10.3.92
Abstract
- BACKGROUND
Nonsteroidal anti-inflammatory drugs (NSAIDs) are widely used to relieve pain, reduce fever and inhibit inflammation. NSAIDs function mainly through inhibition of cyclooxygenase (COX). Growing evidence suggests that NSAIDs also have immunomodulatory effects on T and B cells. Here we examined the effects of NSAIDs on the antigen presenting function of dendritic cells (DCs).
METHODS
DCs were cultured in the presence of aspirin or ibuprofen, and then allowed to phagocytose biodegradable microspheres containing ovalbumin (OVA). After washing and fixing, the efficacy of OVA peptide presentation by DCs was evaluated using OVA-specific CD8 and CD4 T cells.
RESULTS
Aspirin and ibuprofen at high concentrations inhibited both MHC class I and class II-restricted presentation of OVA in DCs. In addition, the DCs generated in the presence of low concentrations of the drugs exhibit a profoundly suppressed capability to present MHC-restricted antigens. Aspirin and ibuprofen did not inhibit the phagocytic activity of DCs, the expression level of total MHC molecules and co-stimulatory molecules on DCs. Ibuprofen rather increased the expression level of total MHC molecules and co-stimulatory molecules on DCs.
CONCLUSION
These results demonstrate that aspirin and ibuprofen inhibit the intracellular processing event of the phagocytosed antigen, and further suggest that prolonged administration of NSAIDs in high doses may impair the capability of DCs to present antigens in asiociation with MHC molecules.
Keyword
MeSH Terms
-
Anti-Inflammatory Agents, Non-Steroidal
Antigen Presentation
Aspirin
B-Lymphocytes
Cyclooxygenase Inhibitors
Dendritic Cells
Fever
Ibuprofen
Inflammation
Microspheres
Ovalbumin
Ovum
Prostaglandin-Endoperoxide Synthases
T-Lymphocytes
Anti-Inflammatory Agents, Non-Steroidal
Aspirin
Cyclooxygenase Inhibitors
Ibuprofen
Ovalbumin
Prostaglandin-Endoperoxide Synthases
Figure
-
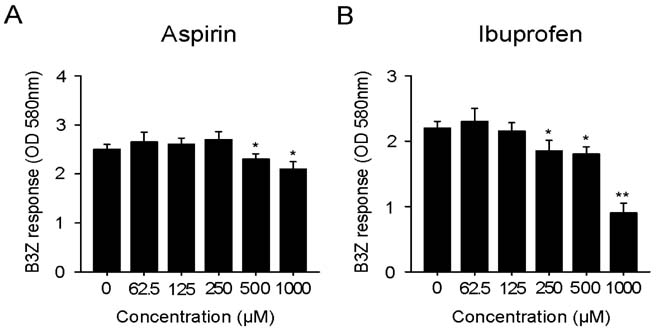
Figure 1 Effects of aspirin (A) and ibuprofen (B) on the cross-presentation of exogenous OVA. DC2.4 cells were incubated with the indicated amounts of the drugs for 18 h, and then added with OVA-microspheres. After 2 h incubation, the cells were washed, fixed, and the amounts of OVA peptides presented on MHC class I molecules were assessed using B3Z cells. The amount of β-galactosidase expressed in B3Z cells was determined by an enzymatic assay using chlorophenolred β-D-galactopyranoside as a substrate. *p<0.05, **p<0.01 compared with untreated control.
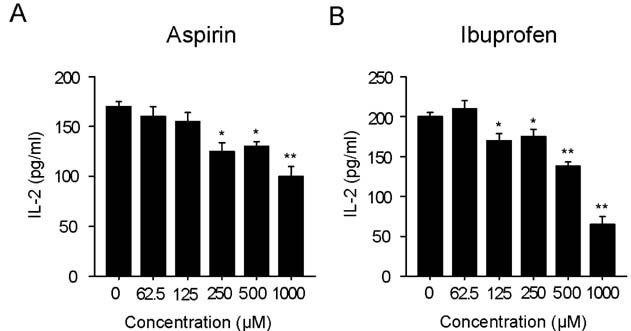
Figure 2 Effects of aspirin and ibuprofen on the class II MHC-restricted presentation of exogenous OVA. Indicated amounts of aspirin or ibuprofen were added to cultures of BM-DCs for 18 h, and then added with OVA-microspheres. After 2 h incubation, the cells were washed, fixed, and the amounts of OVA peptides presented on MHC class II molecules were assessed using OVA-specific CD4 T cell hybridoma, DOBW. The amounts of IL-2 produced from OVA-specific CD4 T cells were assayed by a commercial IL-2 ELISA kit. *p<0.05, **p<0.01 compared with untreated control.
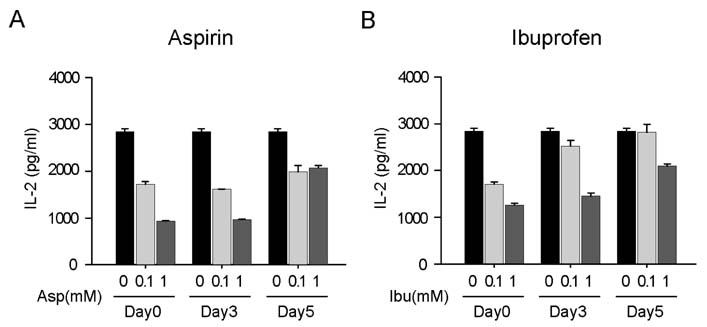
Figure 3 DCs differentiated in the presence of aspirin or ibuprofen are suppressed in MHC class II-restricted antigen presentation capability. Indicated amounts of aspirin or ibuprofen were added to mouse bone marrow cells together with GM-CSF from the initiation of the DC differentiation-inducing culture (day0), at 3 days (Day3), or at 5 days (day5) after the initiation of the cultures. BM-DCs were harvested on day 6 from the initiation of the culture, and the ability to present exogenous antigen in association with MHC class II molecules was ascertained as in Fig. 2.
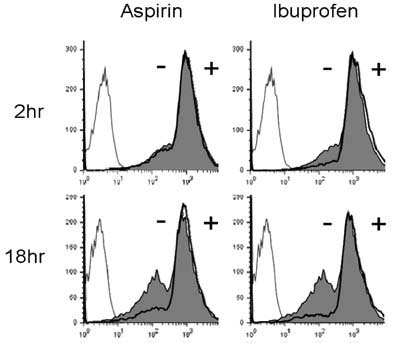
Figure 4 Effects of aspirin and ibuprofen on the phagocytic activity. DC2.4 cells were cultured with aspirin or ibuprofen for 18 h, and then added with microspheres containing both OVA and FITC. After 2 h incubation, unphagocytized microspheres were washed, and the cells were harvested by gentle pipetting, and then analyzed by flow cytometry. Shaded histograms represent the phagocytic activity of DCs cultured in the presence of aspirin or ibuprofen, and thick line histograms represent the phagocytic activity of DCs cultured in the absence of the drugs. DCs that were not incubated with FITC-labeled microspheres are shown as thin line histograms (left).
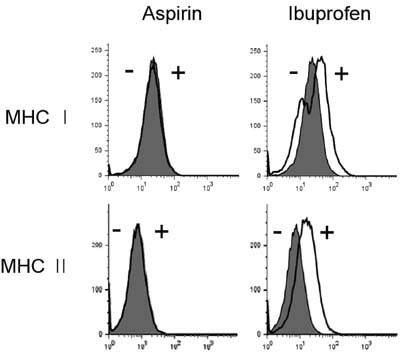
Figure 5 Effects of aspirin and ibuprofen on the expression of MHC molecules. DC2.4 cells were cultured with aspirin or ibuprofen for 18 h, and then the cells were harvested by gentle pipetting. The expression levels of class I and class II MHC molecules were assessed using anti-H-2Kb and anti-I-Ab monoclonal antibodies. Shaded histograms represent the expression levels of H-2Kb and I-Ab molecules in DC2.4 cells cultured in the absence of the drugs. Thick line histograms represent the expression levels of H-2Kb and I-Ab molecules in DC2.4 cells cultured in the presence of the drugs.
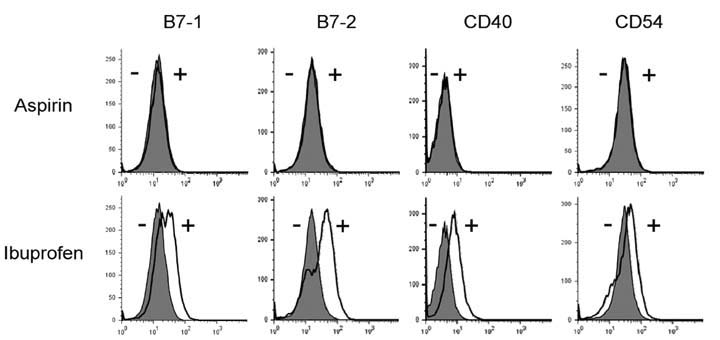
Figure 6 Effects of aspirin and ibuprofen on the expression of co-stimulatory molecules. DC2.4 cells were cultured with aspirin or ibuprofen for 18 h, and then the cells were harvested by gentle pipetting. The expression levels of co-stimulatory molecules were assessed using monoclonal antibodies recognizing mouse cell surface molecules. Shaded histograms represent the expression levels of co-stimulatory molecules in DC2.4 cells cultured in the absence of the drugs. Thick line histograms represent the expression levels of co-stimulatory molecules in DC2.4 cells cultured in the presence of the drugs.
Reference
-
1. Vane JR. Inhibition of prostaglandin synthesis as a mechanism of action for aspirin-like drugs. Nat New Biol. 1971. 231:232–235.
Article2. Smith WL, DeWitt DL, Garavito RM. Cyclooxygenases: structural, cellular, and molecular biology. Annu Rev Biochem. 2000. 69:145–182.
Article3. Smith WL, Dewitt DL. Prostaglandin endoperoxide H synthases-1 and -2. Adv Immunol. 1996. 62:167–215.
Article4. Griswold DE, Adams JL. Constitutive cyclooxygenase (COX-1) and inducible cyclooxygenase (COX-2): rationale for selective inhibition and progress to date. Med Res Rev. 1996. 16:181–206.
Article5. DeWitt DL. Cox-2-selective inhibitors: the new super aspirins. Mol Pharmacol. 1999. 55:625–631.6. Cryer B, Feldman M. Cyclooxygenase-1 and cyclooxygenase-2 selectivity of widely used nonsteroidal anti-inflammatory drugs. Am J Med. 1998. 104:413–421.
Article7. Kazmi SM, Plante RK, Visconti V, Taylor GR, Zhou L, Lau CY. Suppression of NF kappa B activation and NF kappa B-dependent gene expression by tepoxalin, a dual inhibitor of cyclooxygenase and 5-lipoxygenase. J Cell Biochem. 1995. 57:299–310.
Article8. Hall VC, Wolf RE. Effects of tenidap and nonsteroidal anti-inflammatory drugs on the response of cultured human T cells to interleukin 2 in rheumatoid arthritis. J Rheumatol. 1997. 24:1467–1470.9. Iñiguez MA, Punzón C, Fresno M. Induction of cyclooxygenase-2 on activated T lymphocytes: regulation of T cell activation by cyclooxygenase-2 inhibitors. J Immunol. 1999. 163:111–119.10. Kopp E, Ghosh S. Inhibition of NF-kappa B by sodium salicylate and aspirin. Science. 1994. 265:956–959.11. Yin MJ, Yamamoto Y, Gaynor RB. The anti-inflammatory agents aspirin and salicylate inhibit the activity of I(kappa)B kinase-beta. Nature. 1998. 396:77–80.
Article12. Jiang C, Ting AT, Seed B. PPAR-gamma agonists inhibit production of monocyte inflammatory cytokines. Nature. 1998. 391:82–86.
Article13. Bancos S, Bernard MP, Topham DJ, Phipps RP. Ibuprofen and other widely used non-steroidal anti-inflammatory drugs inhibit antibody production in human cells. Cell Immunol. 2009. 258:18–28.
Article14. Paccani SR, Boncristiano M, Ulivieri C, D'Elios MM, Del PG, Baldari CT. Nonsteroidal anti-inflammatory drugs suppress T-cell activation by inhibiting p38 MAPK induction. J Biol Chem. 2002. 277:1509–1513.
Article15. Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K. Immunobiology of dendritic cells. Annu Rev Immunol. 2000. 18:767–811.
Article16. Karttunen J, Sanderson S, Shastri N. Detection of rare antigen-presenting cells by the lacZ T-cell activation assay suggests an expression cloning strategy for T-cell antigens. Proc Natl Acad Sci U S A. 1992. 89:6020–6024.
Article17. Harding CV, Collins DS, Kanagawa O, Unanue ER. Liposome-encapsulated antigens engender lysosomal processing for class II MHC presentation and cytosolic processing for class I presentation. J Immunol. 1991. 147:2860–2863.18. Shen Z, Reznikoff G, Dranoff G, Rock KL. Cloned dendritic cells can present exogenous antigens on both MHC class I and class II molecules. J Immunol. 1997. 158:2723–2730.19. Lee JK, Lee MK, Yun YP, Kim Y, Kim JS, Kim YS, Kim K, Han SS, Lee CK. Acemannan purified from Aloe vera induces phenotypic and functional maturation of immature dendritic cells. Int Immunopharmacol. 2001. 1:1275–1284.
Article20. Lee YH, Lee YR, Im SA, Park SI, Kim KH, Gerelchuluun T, Song S, Kim K, Lee CK. Calcineurin inhibitors block MHC-restricted antigen presentation in vivo. J Immunol. 2007. 179:5711–5716.
Article21. Borthwick GM, Johnson AS, Partington M, Burn J, Wilson R, Arthur HM. Therapeutic levels of aspirin and salicylate directly inhibit a model of angiogenesis through a Cox-independent mechanism. FASEB J. 2006. 20:2009–2016.
Article22. Blain H, Boileau C, Lapicque F, Nédélec E, Loeuille D, Guillaume C, Gaucher A, Jeandel C, Netter P, Jouzeau JY. Limitation of the in vitro whole blood assay for predicting the COX selectivity of NSAIDs in clinical use. Br J Clin Pharmacol. 2002. 53:255–265.
Article23. Heath WR, Carbone FR. Cross-presentation, dendritic cells, tolerance and immunity. Annu Rev Immunol. 2001. 19:47–64.
Article24. Huang AY, Golumbek P, Ahmadzadeh M, Jaffee E, Pardoll D, Levitsky H. Role of bone marrow-derived cells in presenting MHC class I-restricted tumor antigens. Science. 1994. 264:961–965.
Article25. Sigal LJ, Crotty S, Andino R, Rock KL. Cytotoxic T-cell immunity to virus-infected non-haematopoietic cells requires presentation of exogenous antigen. Nature. 1999. 398:77–80.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Immunomodulatory Effects of Hypocrellin A on MHC-restricted Antigen Processing
- Evidence for Direct Inhibition of MHC-Restricted Antigen Processing by Dexamethasone
- Lectins Isolated from Mushroom Fomitella fraxinea Enhance MHC-restricted Exogenous Antigen Presentation
- Inhibition of Major Histocompatibility Complex (MHC)- Restricted Presentation of Exogenous Antigen in Dendritic Cells by Korean Propolis Components
- Vanilloid Receptor 1 Agonists, Capsaicin and Resiniferatoxin, Enhance MHC Class I-restricted Viral Antigen Presentation in Virus-infected Dendritic Cells
