Kidney and Calcium Homeostasis
- Affiliations
-
- 1POSTECH Biotech Center, POSTECH, Pohang, Korea. ujeon@postech.ac.kr
- KMID: 2134808
- DOI: http://doi.org/10.5049/EBP.2008.6.2.68
Abstract
- Plasma calcium concentration is maintained within a narrow range (8.5-10.5 mg/dL) by the coordinated action of parathyroid hormone (PTH), 1,25(OH)2D3, calcitonin, and ionized calcium (iCa2+) itself. The kidney plays a key role in this process by the fine regulation of calcium excretion. More than 95% of filtered calcium is reabsorbed along the renal tubules. In the proximal tubules, 60% of filtered calcium is reabsorbed by passive mechanisms. In the thick ascending limb, 15% of calcium is reabsorbed by paracellular diffusion through paracellin-1 (claudin-16). The calcium sensing receptor (CaSR) in the basolateral membrane of the thick ascending limb senses the change in iCa2+ and inhibits calcium reabsorption independent to PTH and 1,25(OH)2D3. The fine regulation of calcium excretion occurs in the distal convoluted tubules and connecting tubules despite the fact that only 10-15% of filtered calcium is reabsorbed there. Transient receptor potential vanilloid 5 (TRPV5) and 6 (TRPV6) in the apical membrane act as the main portal of entry, calbindin-D28K delivers Ca2+ in the cytoplasm, and then Na2+/Ca2+ exchanger (NCX1) and plasma membrane Ca2+-ATPase in the basolateral membrane serve as an exit. In the cortical collecting duct, TRPV6 is expressed, but the role might be negligible. In addition to PTH and 1,25(OH)2D3, acid-base disturbance, diuretics, and estrogen affect on these calcium channels. Recently, klotho and fibroblast growth factor 23 (FGF23) are suggested as new players in the calcium metabolism. Klotho is exclusively expressed in the kidney and co-localized with TRPV5, NCX1, and calbindin-D28K. Klotho increases calcium reabsorption through trafficking of TRPV5 to the plasma membrane, and also converts FGF receptor to the specific FGF23 receptor. FGF23:klotho complex bound to FGF receptor inhibits 1alpha- hydroxylase of vitamin D, and contributes to calcium reabsorption and phosphate excretion in the kidney.
MeSH Terms
-
Calcitonin
Calcium
Calcium Channels
Calcium-Binding Protein, Vitamin D-Dependent
Cell Membrane
Cytoplasm
Diffusion
Diuretics
Estrogens
Extremities
Fibroblast Growth Factors
Homeostasis
Kidney
Membranes
Parathyroid Hormone
Plasma
Receptors, Calcium-Sensing
Receptors, Fibroblast Growth Factor
TRPV Cation Channels
Vitamin D
Calcitonin
Calcium
Calcium Channels
Calcium-Binding Protein, Vitamin D-Dependent
Diuretics
Estrogens
Fibroblast Growth Factors
Parathyroid Hormone
Receptors, Calcium-Sensing
Receptors, Fibroblast Growth Factor
TRPV Cation Channels
Vitamin D
Figure
-
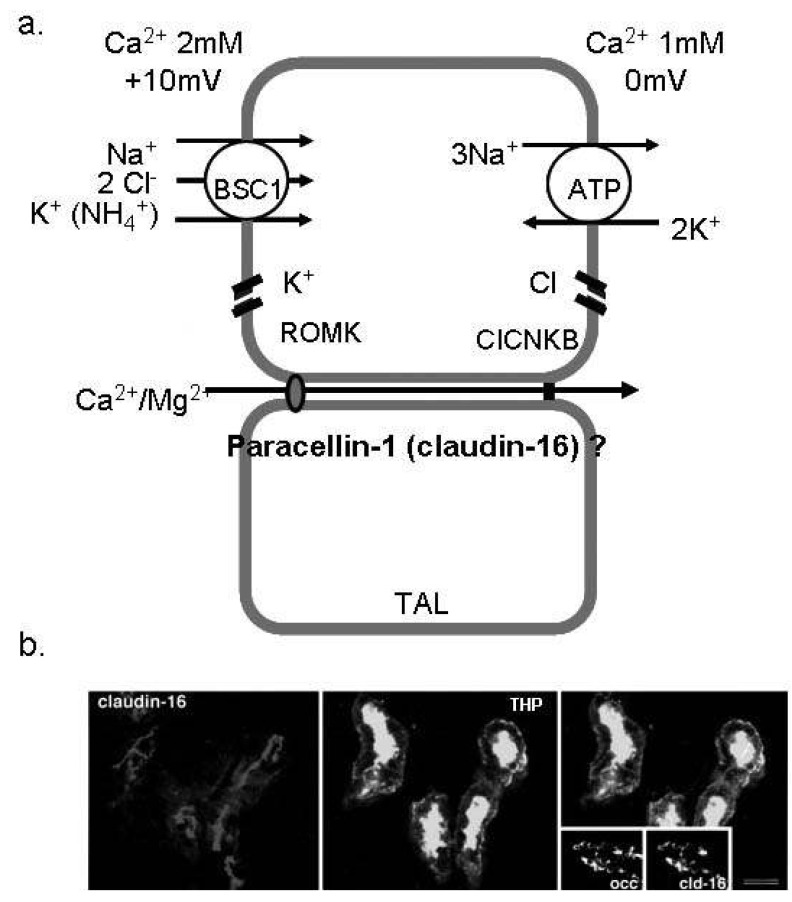
Fig. 1 Ca2+ absorption in the thick ascending limb of Henle (TAL). (a) The schematic view of Ca2+ reabsorption in the TAL. Paracellin-1 (claudin-16) is located in the tight junction of TAL and serves as the paracellular route for divalent cations. (b) Immunofluorescence image for paracellin-1 (claudin-16) in the mouse TAL cells. Paracellin-1 is co-localized with THP (the marker of TAL) and highly expressed in the tight junction1). BSC1, bumetanide sensitive channel; ROMK, renal outer medullary potassium channel; ClCNKB, chloride channel Kb.
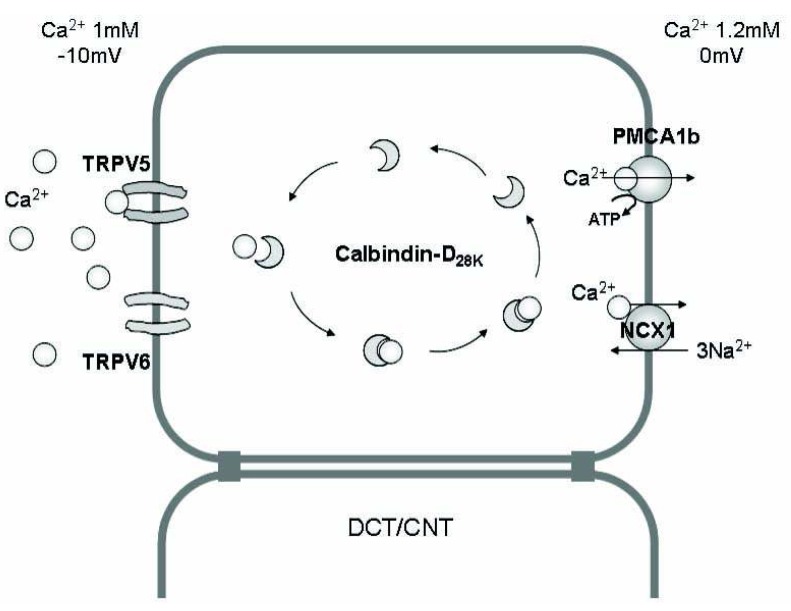
Fig. 2 The mechanism of Ca2+ absorption in the renal epithelium. Transcellular Ca2+ reabsorption in the distal convoluted tubule (DCT) and connecting tubule (CNT) occurs by three steps; (i) entry of Ca2+ through the calcium channels [transient receptor potential vanilloid (TRPV) 5, TRPV6] in the apical membrane, (ii) binding of Ca2+ with calcium-binding protein (calbindin) and diffusion in the cytoplasm (without significant change in the intracellular i[Ca2+]), and (iii) Ca2+ extrusion via an ATP-dependent Ca2+-ATPase (PMCA1b) or an Na2+/Ca2+ exchanger (NCX1) in the basolateral membrane.
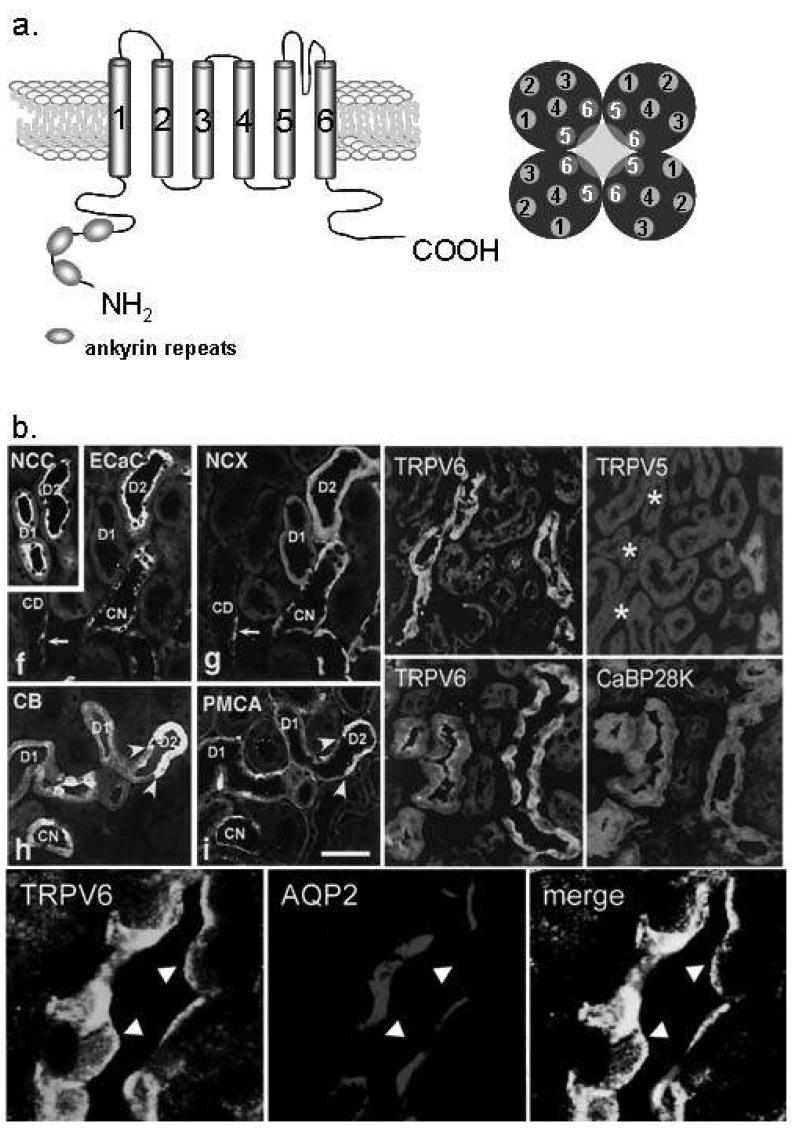
Fig. 3 Ca2+ transport proteins in the distal convoluted tubule (DCT) and connecting tubule (CNT). (a) The molecular structure of transient receptor potential vanilloid (TRPV) 5 and TRPV6. TRPV 5/6 consists of six transmembrane domains and ankyrin repeat at the N-terminal. The functional pore is composed with TM5 and TM6 of the tetramer2). (b) TRPV5 is exclusively expressed in the apical membrane of the DCT and CNT cells (especially DCT-2). TRPV6 is co-localized with TRPV5, but also observed in the collecting duct (the lower panel). Na+/Cl- cotransporter (NCC) is the DCT and CNT marker10, 11). ECaC, epithelial calcilulm channel 1 (TRPV5); NCX, Na2+/Ca2+ exchanger; CaBP28K, calbindin-D28K; AQP2, aquaporin 2.
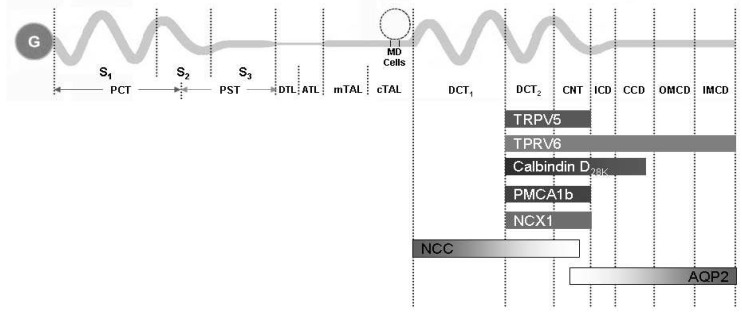
Fig. 4 The summary of the distribution of major renal calcium transport proteins. G, glomerulus; PCT, proximal convoluted tubule; PST, proximal straight tubule; S1, segment 1; S2, segment 2; S3, segment 3; DTL, descending thin limb; ATL, ascending thin limb; mTAL, medullary thick ascending limb; cTAL, cortical thick ascending limb; MD, macula densa; DCT, distal convoluted tubule; CNT, connecting tubule; ICD, initial collecting duct; CCD cortical collecting duct; OMCD, outer medullary collecting duct; IMCD, inner medullary collecting duct; TRPV, transient receptor potential vallinoid; PMCA1b, plasma membrane Ca2+-ATPase; NCX1, Na2+/Ca2+ exchanger; NCC, Na+/Cl- cotransporter; AQP2, aquaporin 2.
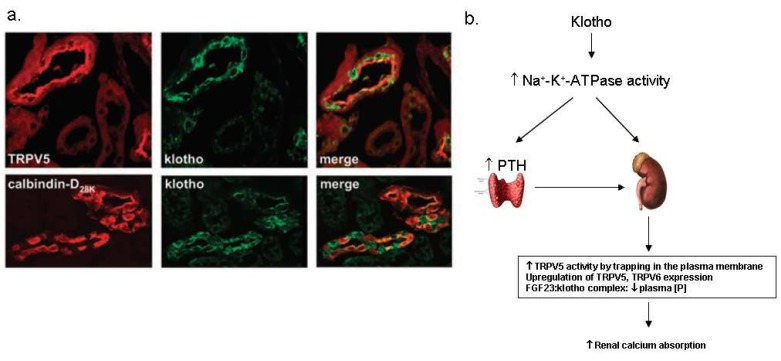
Fig. 5 Klotho in the kidney. (a) The renal distribution of klotho proteins. Klotho is co-expressed with transient receptor potential vallinoid 5 (TRPV5), Na2+/Ca2+ exchanger (NCX1) and calbindin-D28K in the distal convoluted tubule (DCT) and connecting tubule (CNT) 20). (b) The suggested mechanism of the regulation of calcium homeostasis by klotho. PTH, parathyroid hormone; FGF23, fibroblast growth factor 23.
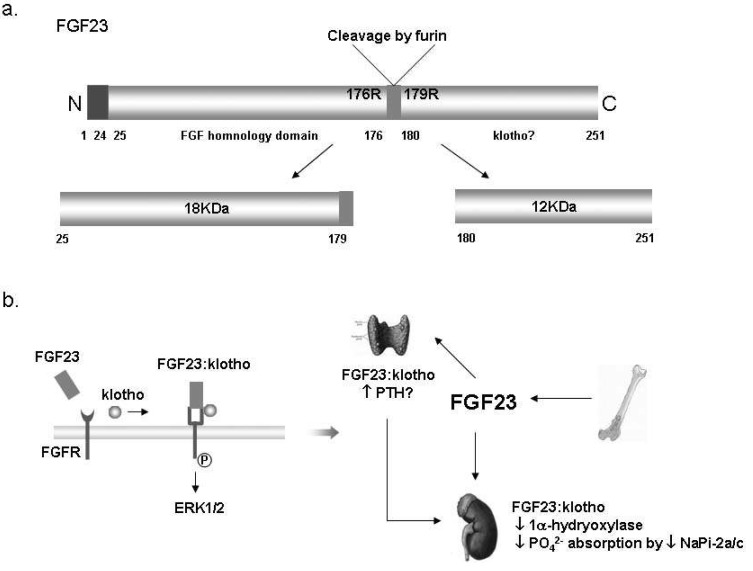
Fig. 6 Fibroblast growth factor 23 (FGF23) in the kidney. (a) The structure of FGF23 proteins. The full length of FGF23 is about 30 kDa, and then inactivated by cleavage to 18 kDa N-terminal fragment and 12 kDa C-terminal fragment24). (b) Suggested mechanism of the FGF23:klotho complex action; Klotho binds to FGF receptor (FGFR) and converts it to a specific receptor for FGF23. Extracellular signalregulated kinase (ERK) 1/2 signaling pathway is activated by binding of the FGF23:klotho complex to FGFR. The FGF23:klotho complex reduces phosphate reabsorption in the proximal tubules via the inhibition of 1α-hydrxylase and NaPi-2a. It might also stimulate parathyroid hormone (PTH) secretion in parathyroid glands. NaPi-2a/c, Type IIa/IIc Na+/Pi contransporter.
Cited by 1 articles
-
Hypercalcemia Associated with Acute Kidney Injury and Metabolic Alkalosis
Jong Hyeok Jeong, Eun Hui Bae
Electrolyte Blood Press. 2010;8(2):92-94. doi: 10.5049/EBP.2010.8.2.92.
Reference
-
1. Mount DB, Yu AS. Brenner BM, editor. Transport of Inorganic Solutes: Sodium, Chloride, Potassium, Magnesium, Calcium, and Phosphate. Brenner & Rector's the kidney. 2008. 8th ed. Philadelphia: p. 185–192.2. Hoenderop JG, Bindels RJ. Epithelial Ca2+ and Mg2+ channels in health and disease. J Am Soc Nephrol. 2005; 16:15–26. PMID: 15574510.3. Nijenhuis T, Renkema KY, Hoenderop JG, Bindels RJ. Acid-base status determines the renal expression of Ca2+ and Mg2+ transport proteins. J Am Soc Nephrol. 2006; 17:617–626. PMID: 16421227.4. Lee CT, Chen HC, Lai LW, Yong KC, Lien YH. Effects of furosemide on renal calcium handling. Am J Physiol Renal Physiol. 2007; 293:F1231–F1237. PMID: 17652376.
Article5. Nijenhuis T, Hoenderop JG, Loffing J, van der Kemp AW, van Os CH, Bindels RJ. Thiazide-induced hypocalciuria is accompanied by a decreased expression of Ca2+ transport proteins in kidney. Kidney Int. 2003; 64:555–564. PMID: 12846750.6. Kiuchi-Saishin Y, Gotoh S, Furuse M, Takasuga A, Tano Y, Tsukita S. Differential expression patterns of claudins, tight junction membrane proteins, in mouse nephron segments. J Am Soc Nephrol. 2002; 13:875–886. PMID: 11912246.
Article7. Ward DT, Riccardi D. Renal physiology of the extracellular calcium-sensing receptor. Pflugers Arch. 2002; 445:169–176. PMID: 12457237.
Article8. Attie MF, Gill JR Jr, Stock JL, Spiegel AM, Downs RW Jr, Levine MA, et al. Urinary calcium excretion in familial hypocalciuric hypercalcemia. Persistence of relative hypocalciuria after induction of hypoparathyroidism. J Clin Invest. 1983; 72:667–676. PMID: 6874959.
Article9. van de Graaf SF, Hoenderop JG, Bindels RJ. Regulation of TRPV5 and TRPV6 by associated proteins. Am J Physiol Renal Physiol. 2006; 290:F1295–F1302. PMID: 16682485.
Article10. Hoenderop JG, Muller D, Van Der Kemp AW, Hartog A, Suzuki M, Ishibashi K, et al. Calcitriol controls the epithelial calcium channel in kidney. J Am Soc Nephrol. 2001; 12:1342–1349. PMID: 11423563.
Article11. Nijenhuis T, Hoenderop JG, van der Kemp AW, Bindels RJ. Localization and regulation of the epithelial Ca2+ channel TRPV6 in the kidney. J Am Soc Nephrol. 2003; 14:2731–2740. PMID: 14569082.12. Hoenderop JG, van Leeuwen JP, van der Eerden BC, Kersten FF, van der Kemp AW, Merillat AM, et al. Renal Ca2+ wasting, hyperabsorption, and reduced bone thickness in mice lacking TRPV5. J Clin Invest. 2003; 112:1906–1914. PMID: 14679186.13. Hoenderop JG, Bindels RJ. Calciotropic and magnesiotropic TRP channels. Physiology (Bethesda). 2008; 23:32–40. PMID: 18268363.
Article14. Nijenhuis T, Vallon V, van der Kemp AW, Loffing J, Hoenderop JG, Bindels RJ. Enhanced passive Ca2+ reabsorption and reduced Mg2+ channel abundance explains thiazide-induced hypocalciuria and hypomagnesemia. J Clin Invest. 2005; 115:1651–1658. PMID: 15902302.15. Lee CT, Shang S, Lai LW, Yong KC, Lien YH. Effect of thiazide on renal gene expression of apical calcium channels and calbindins. Am J Physiol Renal Physiol. 2004; 287:F1164–F1170. PMID: 15265769.
Article16. Jang HR, Lee JW, Heo NJ, Lee JH, Oh YK, Na KY, et al. Effects of thiazide on the expression of transient receptor potential vanilloid 5 and calbindin-D28K in a hypercalciuria rat model [Abstract]. J Am Soc Nephrol. 2006; 17:355A.17. Lambers TT, Mahieu F, Oancea E, Hoofd L, de Lange F, Mensenkamp AR, et al. Calbindin-D28K dynamically controls TRPV5-mediated Ca2+ transport. EMBO J. 2006; 25:2978–2988. PMID: 16763551.18. Kuro-o M, Matsumura Y, Aizawa H, Kawaguchi H, Suga T, Utsugi T, et al. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature. 1997; 390:45–51. PMID: 9363890.
Article19. Imura A, Tsuji Y, Murata M, Maeda R, Kubota K, Iwano A, et al. alpha-Klotho as a regulator of calcium homeostasis. Science. 2007; 316:1615–1618. PMID: 17569864.20. Chang Q, Hoefs S, van der Kemp AW, Topala CN, Bindels RJ, Hoenderop JG. The beta-glucuronidase klotho hydrolyzes and activates the TRPV5 channel. Science. 2005; 310:490–493. PMID: 16239475.21. Nabeshima Y, Imura H. alpha-Klotho: a regulator that integrates calcium homeostasis. Am J Nephrol. 2008; 28:455–464. PMID: 18160815.22. Urakawa I, Yamazaki Y, Shimada T, Iijima K, Hasegawa H, Okawa K, et al. Klotho converts canonical FGF receptor into a specific receptor for FGF23. Nature. 2006; 444:770–774. PMID: 17086194.
Article23. Yamashita T, Yoshioka M, Itoh N. Identification of a novel fibroblast growth factor, FGF-23, preferentially expressed in the ventrolateral thalamic nucleus of the brain. Biochem Biophys Res Commun. 2000; 277:494–498. PMID: 11032749.
Article24. Razzaque MS, Lanske B. The emerging role of the fibroblast growth factor-23-klotho axis in renal regulation of phosphate homeostasis. J Endocrinol. 2007; 194:1–10. PMID: 17592015.
Article
