Beta-Defensin 124 Is Required for Efficient Innate Immune Responses in Prostate Epithelial RWPE-1 Cells
- Affiliations
-
- 1Research Institute for Translational System Biomics, Chung-Ang University College of Medicine, Seoul, Korea. uromyung@cau.ac.kr
- 2Department of Urology, Hallym University Dontan Sacred Heart Hospital, Hwaseong, Korea.
- 3Department of Urology, Chung-Ang University College of Medicine, Seoul, Korea.
- KMID: 1885622
- DOI: http://doi.org/10.4111/kju.2014.55.6.417
Abstract
- PURPOSE
The present study aimed to determine the role played by beta-defensin 124 (DEFB124) in the innate immunity of prostate epithelial RWPE-1 cells during bacterial infection.
MATERIALS AND METHODS
The expression of DEFB124 was examined by quantitative real-time polymerase chain reaction (PCR), Western blotting, and immunocytochemistry. Enzyme-linked immunosorbent assays and quantitative real-time PCR were performed to determine the production of cytokines and chemokines. Western blotting and chromatin immunoprecipitation studies were performed to assess the interaction between DEFB124 and nuclear factor-kappa B (NF-kappaB) in peptidoglycan (PGN)-stimulated RWPE-1 cells. By chemotaxis assay, we assessed the effect of DEFB124 on the migration of monocytes.
RESULTS
Exposure to PGN induced DEFB124 upregulation and NF-kappaB activation through IkappaBalpha phosphorylation and IkappaBalpha degradation. Bay11-7082, an NF-kappaB inhibitor, blocked PGN-induced DEFB124 production. Also, NF-kappaB was shown to be a direct regulator and to directly bind to the -3.14 kb site of the DEFB124 promoter in PGN-treated human prostate epithelial RWPE-1 cells. When DEFB124 was overexpressed in RWPE-1 cells, interestingly, the production of cytokines (interleukin [IL] 6 and IL-12) and chemokines (CCL5, CCL22, and CXCL8) was significantly increased. These DEFB124-upregulated RWPE-1 cells markedly induced chemotactic activity for THP-1 monocytes.
CONCLUSIONS
Taken together, these results provide strong evidence for the first time that increased DEFB124 expression via NF-kappaB activation in PGN-exposed RWPE-1 cells enhances the production of cytokines and chemokines, which may contribute to an efficient innate immune defense.
Keyword
MeSH Terms
-
Bacterial Infections
Blotting, Western
Chemokines
Chemotaxis
Chromatin Immunoprecipitation
Cytokines
Defensins
Enzyme-Linked Immunosorbent Assay
Humans
Immunity, Innate*
Immunohistochemistry
Monocytes
NF-kappa B
Peptidoglycan
Phosphorylation
Prostate*
Real-Time Polymerase Chain Reaction
Up-Regulation
Chemokines
Cytokines
Defensins
NF-kappa B
Peptidoglycan
Figure
-

FIG. 1 Peptidoglycan (PGN) induces gene expression of β-defensin 124 (DEFB124) through nuclear factor-kappa B (NF-κB) activation in RWPE-1 cells. (A) PGN induces DEFB124 mRNA expression. The RWPE-1 cells were treated with PGN (10 µg/mL) for the indicated times and DEFB124 expression was determined by reverse transcription-polymerase chain reaction. (B) PGN leads to NF-κB activation in RWPE-1 cells. The cells were stimulated with 10 µg/mL of PGN, and protein was extracted at the indicated time points. The phosphorylation and ubiquitination of IκBα were examined by western blot. ACTB was used as an internal control. (C) The activation of NF-κB is required for the upregulation of PGN-induced DEFB124. The RWPE-1 cells were treated with PGN or NF-κB inhibitor Bay11-7082 (10 µM), and the concentration of secreted DEFB124 protein was measured in the culture supernatant by enzyme-linked immunosorbent assay. Concentrations are picograms of protein per mL, and data are the mean results of three distinct experiments. Different letters indicate significant differences at p<0.0001. (D) PGN induces DEFB124 production through NF-κB activation. The RWPE-1 cells were treated with PGN or Bay11-7082, and reduced production of PGN-induced DEFB124 protein was evaluated by immunocytochemistry. RWPE-1 cells were fixed in 4% paraformaldehyde, stained with anti-DEFB124 antibody (green), and counterstained with PI (red) for DNA staining. (E) NF-κB interacts with DEFB124 chromatin structure. NF-κB directly binds at the -3.14 Kb site of DEFB124 in PGN-stimulated RWPE-1 cells. Results, normalized for the input DNA and expressed as the relative enrichment of immunoprecipitated PGN-treated RWPE-1 cells compared to the untreated control RWPE-1 cells, are the means of three independent experiments. The site difference from the consensus NF-κB binding sequences (CBS) are indicated by asterisks. CBS, GGGRNNYYCC; R, purine; Y, pyrimidine. Different letters indicate significant differences at p<0.0001.
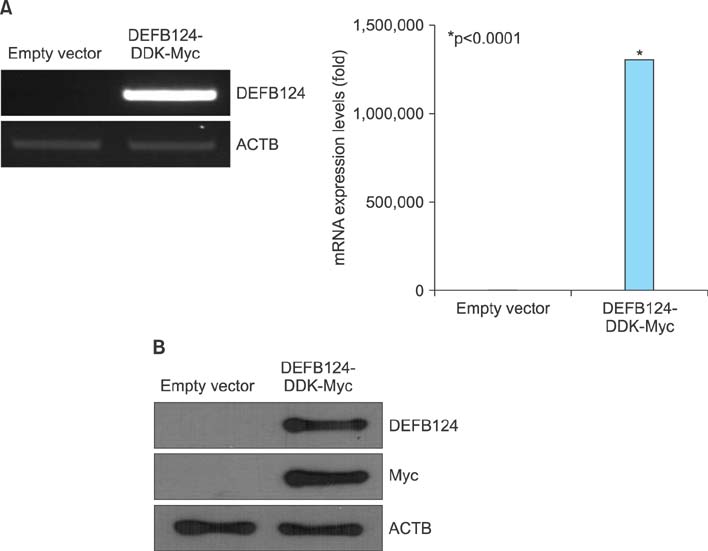
FIG. 2 Overexpression of β-defensin 124 (DEFB124) in RWPE-1 cells. (A) DEFB124 mRNA overexpression. The RWPE-1 cells were transfected with DEFB124-DDK-Myc vector or empty vector, and DEFB124 mRNA expression was determined by reverse transcription-polymerase chain reaction (left) and quantitative real-time polymerase chain reaction (right). ACTB was used as an internal control. Asterisk represents statistical significance at p<0.0001. (B) Upregulation of DEFB124 protein. DEFB124 protein was evaluated by western blot by using antibodies against DEFB124 and Myc. ACTB was used as an internal control.

FIG. 3 Beta-Defensin 124 (DEFB124) upregulation induces increased production of cytokines and chemokines. DEFB124 promotes mRNA expression for cytokines (A) and chemokines (B) in DEFB124-induced RWPE-1 cells. The mRNA expression of these genes was determined by using quantitative real-time polymerase chain reaction. Relative expression levels of each gene were calculated from cycle threshold values and were normalized with ACTB, and the expression ratio was calculated against the expression of each gene in the empty vector-transfected RWPE-1 cells. Experiments were repeated at least three times, and data are expressed as the mean±standard error of the mean (SEM). Asterisks, * and **, represent statistical significance at p<0.05 and p<0.01, respectively. (C) DEFB124 is required for cytokine and chemokine production. Supernatants from empty vector- or DEFB124-DDK-Myctransfected RWPE-1 cells were collected. The concentrations of cytokines and chemokines in supernatant were measured by multianalyte enzyme-linked immunosorbent assay (ELISA). The diagram shows mean ELISA absorbance values (450 nm) for triplicates with the error bars representing the SEM. Asterisks (*, **, and ***) represent statistical significance at p<0.02, p<0.005, and p<0.0001, respectively. IL, interleukin.
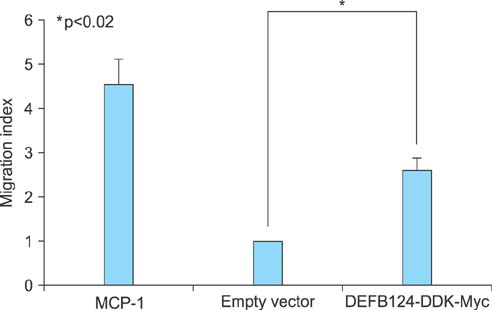
FIG. 4 Beta-Defensin 124 (DEFB124)-mediated upregulation of cytokines and chemokines promote chemotactic response of THP-1 monocytes. DEFB124- or DEFB124-mediated cytokines and chemokines induce chemotaxis for THP-1 monocytes. THP-1 cells (1×106 cells) were added to the upper chamber, and the lower chamber contained supernatants from either the empty vector- or DEFB124-DDK-Myc-transfected RWPE-1 cells. The results are presented as a migration index denoting the fold increase of cell migration over the empty vector. Monocyte chemoattractant protein-1 (MCP-1) (100 ng/mL) was used as a positive control. Results are representative of three independent experiments. Asterisk represents statistical significance at p<0.02.
Reference
-
1. Boman HG. Peptide antibiotics and their role in innate immunity. Annu Rev Immunol. 1995; 13:61–92.2. Nizet V. Antimicrobial peptide resistance mechanisms of human bacterial pathogens. Curr Issues Mol Biol. 2006; 8:11–26.3. Izadpanah A, Gallo RL. Antimicrobial peptides. J Am Acad Dermatol. 2005; 52(3 Pt 1):381–390.4. Nguyen LT, Haney EF, Vogel HJ. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011; 29:464–472.5. Ganz T. The role of antimicrobial peptides in innate immunity. Integr Comp Biol. 2003; 43:300–304.6. Ganz T. Defensins: antimicrobial peptides of vertebrates. C R Biol. 2004; 327:539–549.7. Tang YQ, Yuan J, Miller CJ, Selsted ME. Isolation, characterization, cDNA cloning, and antimicrobial properties of two distinct subfamilies of alpha-defensins from rhesus macaque leukocytes. Infect Immun. 1999; 67:6139–6144.8. Lai Y, Gallo RL. AMPed up immunity: how antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009; 30:131–141.9. Ganz T. Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol. 2003; 3:710–720.10. Wei G, de Leeuw E, Pazgier M, Yuan W, Zou G, Wang J, et al. Through the looking glass, mechanistic insights from enantiomeric human defensins. J Biol Chem. 2009; 284:29180–29192.11. Schutte BC, Mitros JP, Bartlett JA, Walters JD, Jia HP, Welsh MJ, et al. Discovery of five conserved beta -defensin gene clusters using a computational search strategy. Proc Natl Acad Sci U S A. 2002; 99:2129–2133.12. Bajaj-Elliott M, Fedeli P, Smith GV, Domizio P, Maher L, Ali RS, et al. Modulation of host antimicrobial peptide (beta-defensins 1 and 2) expression during gastritis. Gut. 2002; 51:356–361.13. Motzkus D, Schulz-Maronde S, Heitland A, Schulz A, Forssmann WG, Jubner M, et al. The novel beta-defensin DEFB123 prevents lipopolysaccharide-mediated effects in vitro and in vivo. FASEB J. 2006; 20:1701–1702.14. Bonizzi G, Karin M. The two NF-kappaB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 2004; 25:280–288.15. Kim HJ, Jung JR, Kim HJ, Lee SY, Chang IH, Lee TJ, et al. Expression of human β-defensin-2 in the prostate. BJU Int. 2011; 107:144–149.16. Andresen L, Jorgensen VL, Perner A, Hansen A, Eugen-Olsen J, Rask-Madsen J. Activation of nuclear factor kappaB in colonic mucosa from patients with collagenous and ulcerative colitis. Gut. 2005; 54:503–509.17. Lawrence T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb Perspect Biol. 2009; 1:a001651.18. Wehkamp J, Harder J, Wehkamp K, Wehkamp-von Meissner B, Schlee M, Enders C, et al. NF-kappaB- and AP-1-mediated induction of human beta defensin-2 in intestinal epithelial cells by Escherichia coli Nissle 1917: a novel effect of a probiotic bacterium. Infect Immun. 2004; 72:5750–5758.19. Steubesand N, Kiehne K, Brunke G, Pahl R, Reiss K, Herzig KH, et al. The expression of the beta-defensins hBD-2 and hBD-3 is differentially regulated by NF-kappaB and MAPK/AP-1 pathways in an in vitro model of Candida esophagitis. BMC Immunol. 2009; 10:36.20. Mburu YK, Abe K, Ferris LK, Sarkar SN, Ferris RL. Human β-defensin 3 promotes NF-κB-mediated CCR7 expression and anti-apoptotic signals in squamous cell carcinoma of the head and neck. Carcinogenesis. 2011; 32:168–174.21. Watford WT, Moriguchi M, Morinobu A, O'Shea JJ. The biology of IL-12: coordinating innate and adaptive immune responses. Cytokine Growth Factor Rev. 2003; 14:361–368.22. Jones LL, Vignali DA. Molecular interactions within the IL-6/IL-12 cytokine/receptor superfamily. Immunol Res. 2011; 51:5–14.23. Luster AD. The role of chemokines in linking innate and adaptive immunity. Curr Opin Immunol. 2002; 14:129–135.24. Robertson MJ. Role of chemokines in the biology of natural killer cells. J Leukoc Biol. 2002; 71:173–183.25. Amerio P, Frezzolini A, Abeni D, Teofoli P, Girardelli CR, De Pita O, et al. Increased IL-18 in patients with systemic lupus erythematosus: relations with Th-1, Th-2, pro-inflammatory cytokines and disease activity. IL-18 is a marker of disease activity but does not correlate with pro-inflammatory cytokines. Clin Exp Rheumatol. 2002; 20:535–538.26. Maczynska I, Millo B, Ratajczak-Stefanska V, Maleszka R, Szych Z, Kurpisz M, et al. Proinflammatory cytokine (IL-1beta, IL-6, IL-12, IL-18 and TNF-alpha) levels in sera of patients with subacute cutaneous lupus erythematosus (SCLE). Immunol Lett. 2006; 102:79–82.27. Berin MC, Dwinell MB, Eckmann L, Kagnoff MF. Production of MDC/CCL22 by human intestinal epithelial cells. Am J Physiol Gastrointest Liver Physiol. 2001; 280:G1217–G1226.28. Murdoch C, Read RC, Zhang Q, Finn A. Choline-binding protein A of Streptococcus pneumoniae elicits chemokine production and expression of intercellular adhesion molecule 1 (CD54) by human alveolar epithelial cells. J Infect Dis. 2002; 186:1253–1260.29. Kim CH, Broxmeyer HE. Chemokines: signal lamps for trafficking of T and B cells for development and effector function. J Leukoc Biol. 1999; 65:6–15.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Gene Regulatory Regions Required for beta Defensin-2 Up-Regulation by Interleukin-1alpha in the Human Middle Ear Epithelial Cell Line
- Expression of beta Defensins in the Human Middle Ear Mucosa
- Expression of Antimicrobial Defensin Peptides of the Human Nasal Mucosa
- A Dynamic Interplay of Innate Immune Responses During Urinary Tract Infection
- The Innate Immune Responses in Pathogenesis of Chronic Rhinosinusitis
