Relationship Between the Extent of Chromosomal Losses and the Pattern of CpG Methylation in Gastric Carcinomas
- Affiliations
-
- 1Department of Microbiology, College of Medicine, The Catholic University of Korea, Seoul, Korea. rhyumung@catholic.ac.kr
- 2Department of Clinical Pathology, College of Medicine, The Catholic University of Korea, Seoul, Korea.
- 3Department of Internal Medicine, College of Medicine, The Catholic University of Korea, Seoul, Korea.
- KMID: 1781762
- DOI: http://doi.org/10.3346/jkms.2005.20.5.790
Abstract
- The extent of unilateral chromosomal losses and the presence of microsatellite instability (MSI) have been classified into high-risk (high- and baseline-level loss) and low-risk (low-level loss and MSI) stem-line genotypes in gastric carcinomas. A unilateral genome-dosage reduction might stimulate compensation mechanism, which maintains the genomic dosage via CpG hypomethylation. A total of 120 tumor sites from 40 gastric carcinomas were examined by chromosomal loss analysis using 40 microsatellite markers on 8 chromosomes and methylation analysis in the 13 CpG (island/non-island) regions near the 10 genes using the bisulfite-modified DNAs. The high-level-loss tumor (four or more losses) showed a tendency toward unmethylation in the Maspin, CAGE, MAGE-A2 and RABGEF1 genes, and the other microsatellite-genotype (three or fewer losses and MSI) toward methylation in the p16, hMLH1, RASSF1A, and Cyclin D2 genes (p<0.05). The non-island CpGs of the p16 and hMLH1 genes were hypomethylated in the high-level-loss and hypermethylated in the non-high-level-loss sites (p<0.05). Consequently, hypomethylation changes were related to a high-level loss, whereas the hypermethylation changes were accompanied by a baseline-level loss, a low-level loss, or a MSI. This indicates that hypomethylation compensates the chromosomal losses in the process of tumor progression.
Keyword
MeSH Terms
-
Chromosome Aberrations/*statistics and numerical data
Chromosome Mapping/*methods
CpG Islands/*genetics
*DNA Methylation
DNA Mutational Analysis/methods
France/epidemiology
Genetic Predisposition to Disease/epidemiology/genetics
Genetic Screening/methods
Genomic Instability/genetics
Humans
Incidence
Korea/epidemiology
Microsatellite Repeats/genetics
Polymorphism, Genetic
Research Support, Non-U.S. Gov't
Risk Assessment/*methods
Risk Factors
Statistics
Stomach Neoplasms/*enzymology/*genetics
Figure
-
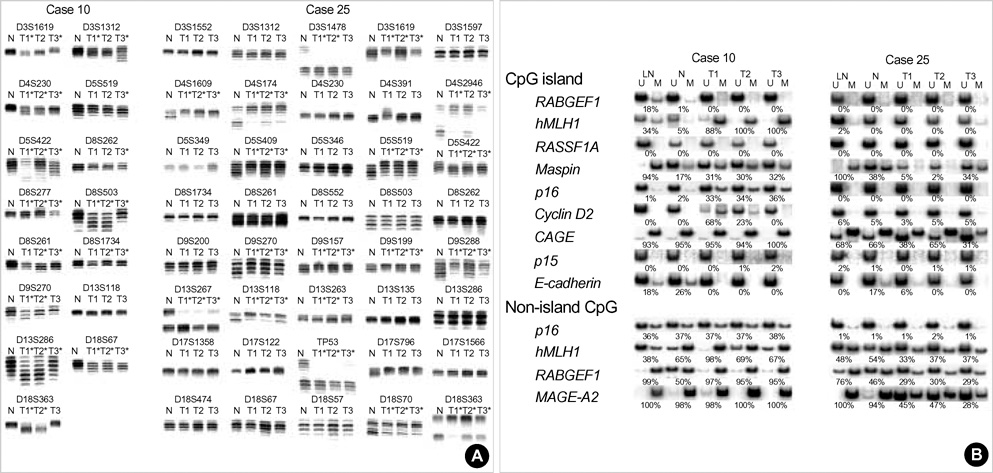
Fig. 1 Representative autoradiographs of the multifocal microsatellite and methylation analyses. (A) Case 10 shows a high-frequency microsatellite instability (MSI) at more than 40% of the 15 homozygous markers in the three tumor sites (T1, 67%; T2, 73%; T3, 80%). Case 25 had a high-level loss of heterozygosity (LOH) involving chromosomes 3p, 4p, 5q, 9p, 13q, 17p, and 18q commonly shared by the three tumor sites. Normal (N) and the corresponding tumor (T) DNAs are indicated above each allelic band. The asterisk indicates a MSI or LOH. (B) Normal lymph node (LN), nontumoral gastric mucosa (N), and multiple tumor sites (T1, T2, and T3) have a broad range of methylated and unmethylated amplification from the bisulfite-modified DNAs. The lanes marked by U and M indicate the PCR bands of the unmethylation and methylation primer sets, respectively. The methylated and unmethylated proportion of the Maspin CpG islands and the RABGEF1 non-island CpGs was obviously different between the normal lymph node and the gastric mucosa of case 10 and 25. Case 10 with a MSI shows diverse increases in the methylated proportion of the CpG islands of the hMLH1, Maspin, p16, and Cyclin D2 genes as well as the non-island CpGs of the hMLH1 and RABGEF1 genes. In case 25 with a high-level chromosomal loss, the unmethylated proportion of the CpG islands of the Maspin and CAGE genes and the non-island CpGs of the hMLH1, RABGEF1 and MAGE-A2 genes are increased in a similar or dissimilar grade. The percentage of methylated CpGs in the template DNA calculated based on the standard MSP curve (Fig. 2) is indicated below the lanes. The results of microsatellite and methylation analyses are summarized in Tables 2 and 5, respectively.
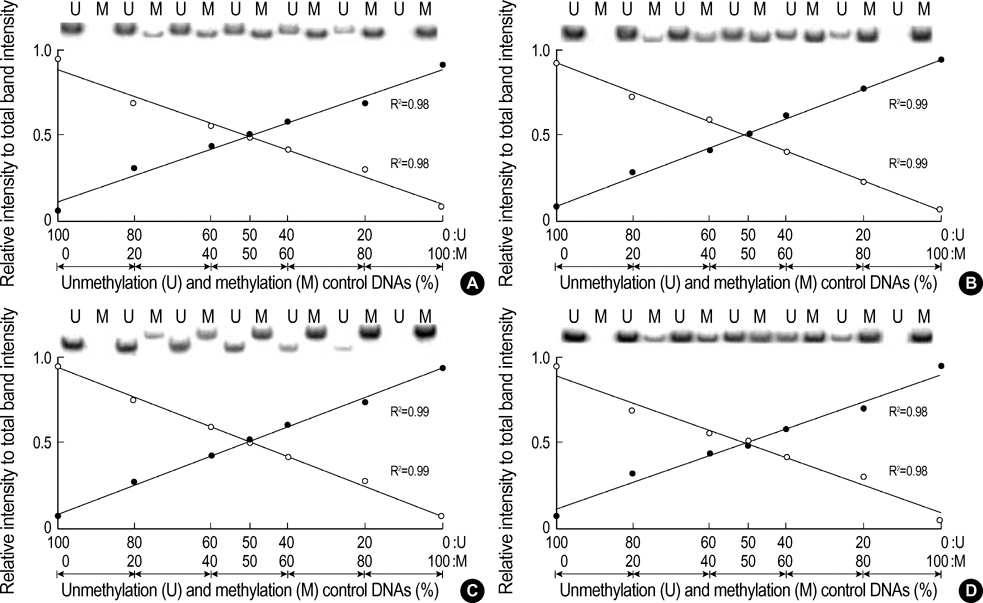
Fig. 2 Standard curves for the methylated and unmethylated MSP amplification. The genomic DNA universally methylated by DNA methylase and the genome-wide-PCR DNA amplified by a universal primer were used as the positive controls for the methylation and unmethylation DNAs, respectively. The variable mixtures of the two opposite control DNAs based on their PCR intensity of 20 ng/µL were amplified using a set of MSP primers. The band intensities of the methylation (M, closed circle) and unmethylation (U, open circle) primer sets for the CpG islands of the hMLH1 (A), Maspin (B), and CAGE (C) genes and the non-island CpGs of hMLH1 gene (D) are plotted along with the relative proportion against the total band intensity. The amplification intensity ratio of each primer set is linearly increased with the increasing percentage of the corresponding control DNA in the MSP template-primer mixtures. The methylation-unmethylation proportion of the template DNA was distinguished into five grades (0-20% methylation, 21-40% methylation, 41-60% methylation, 61-80% methylation, and 81-100% methylation) based on the reciprocal curves of the methylation and unmethylation bands. The band intensity ratio of each MSP primer set is listed in Table 1.
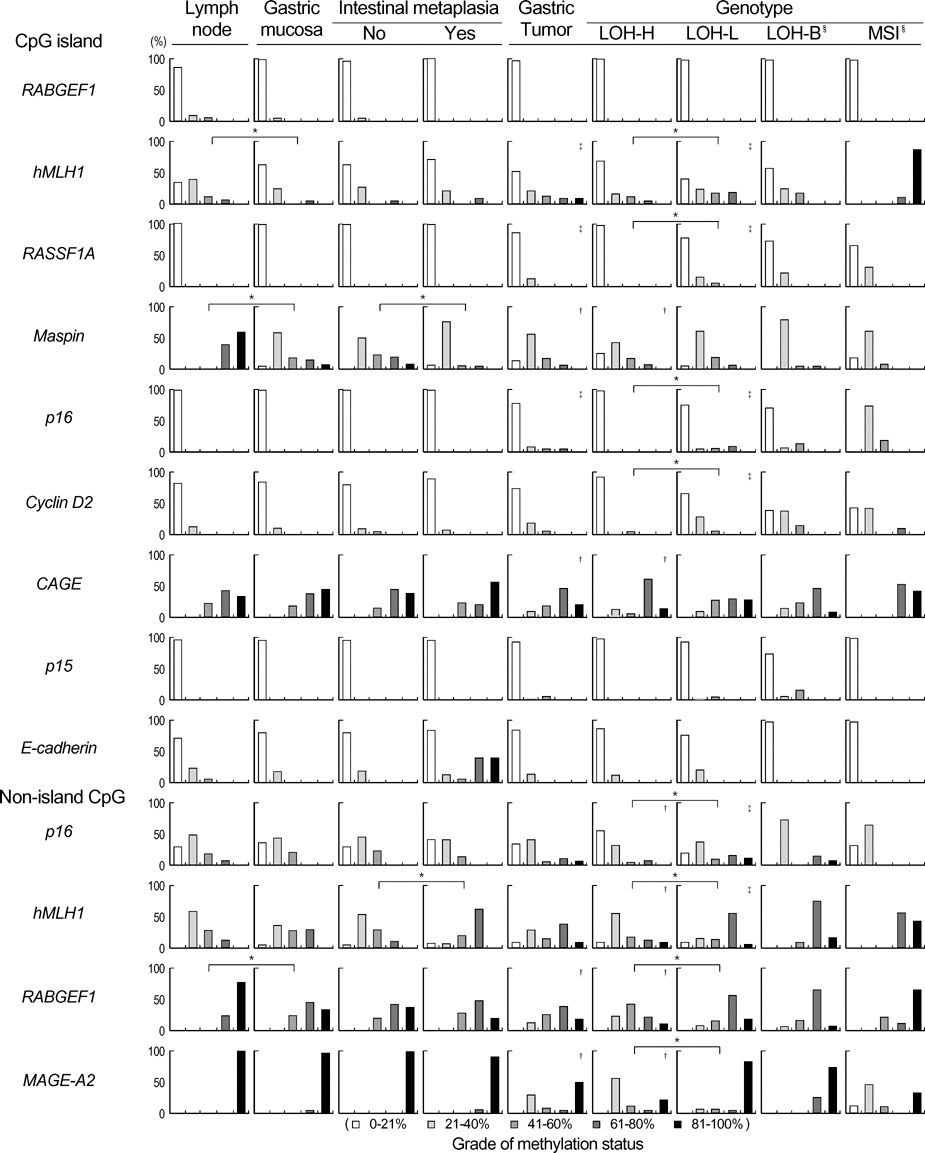
Fig. 3 Methylation profiles of the CpG islands and non-island CpGs in the nontumoral and tumoral tissues. The nontumoral gastric mucosa is grouped according to the presence and absence of the intestinal metaplasia. The tumor tissues are grouped into four microsatellite genotypes. The classification of the microsatellite genotypes is detailed in the "Materials and Methods" section. The CpG methylation status in each tissue group is given as a percentage of five grades. The p-value was calculated using the Mann-Whitney U test. There were *significant differences in methylation density between the lymph node and gastric mucosa, between the presence and absence of intestinal metaplasia, and between the LOH-H and LOH-L groups. The heterogeneous tumor sites showed significant †decreased methylation and ‡increased methylation density compared with the gastric mucosa. §Statistical comparison was not shown due to the small number of LOH-B and MSI cases. All p-values are listed in the Table 7.
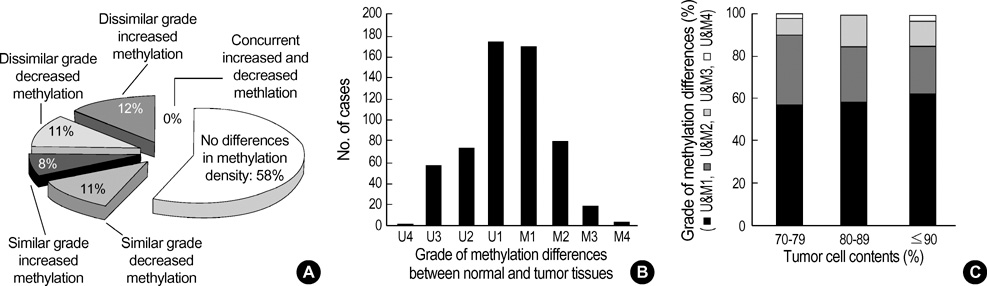
Fig. 4 Methylation heterogeneity on gastric carcinomas detected by multifocal analysis. The methylation status of the 13 CpG regions is detailed in Table 6. (A) The grade and pattern of the methylation differences in a given CpG region in different tumor sites from a given gastric carcinoma were compared. (B) The methylation differences detected in individual tumor sites are shown separately according to the grade of the methylation differences. (C) The grades of the methylation differences given as percentage were compared between different tumor-cell-content groups. The differences in the methylated-unmethylated proportion between the paired normal and heterogeneous tumor sites were graded U1-U4 decreased methylation differences and M1-M4 increased methylation differences.

Fig. 5 Heterogeneous methylation profiles of the p16, hMLH1, and MAGE-A2 CpG regions. PCR products common for methylated and unmethylated CpGs were cloned and ten clones were randomly selected. The individual and pooled clones were amplified by the MSP primer sets (A). The individual PCR clones generated heterogeneous methylation densities in the MSP analysis. The pooled PCR clones showed similar MSP patterns with those of the corresponding genomic DNAs listed in Table 6. Five PCR clones from each genomic DNA generating different or no MSP band intensities were sequenced (B). PCR primers specific for the unmethylation ( ) and methylation (▬), and common (▭) for the methylation and unmethylation are indicated by the horizontal bars below the vertical lines marking the individual CpG sites. Individual CpG sites of each PCR clone are indicated as circles in a clone-numbered row. A closed circle marks a methylated CpG and an open circle marks an unmethylated CpG. The methylation composition of the PCR clones was heterogeneous in similarity with a broad range of methylated and unmethylated band intensities. One or two primer-template sequence mismatches produced weak band intensities. Multiple sequence mismatches in the primer regions of the hMLH1 CpG islands (normal clone 6 and tumor clone 4 in case 43) produced none of the methylation and unmethylation bands.
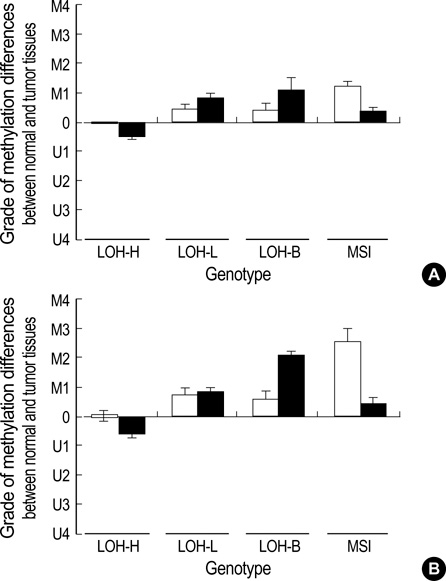
Fig. 6 Pattern of methylation differences in the CpG island (□) and non-island CpG (■) area of the p16 (A) and hMLH1 (B) genes. The gastric carcinoma was classified as four microsatellite genotypes. The classification of the microsatellite genotypes is detailed in the "Materials and Methods" section. The decreased methylation differences in the LOH-H cases and the increased methylation differences in the LOH-L, LOH-B, and MSI cases were observed in the methylation gradient region between the CpG islands and non-island CpG area of the same gene. The increased methylation (M1-4) and decreased methylation (U1-4) differences are indicated by a difference in methylation grade between the gastric mucosa and the gastric carcinoma. The bar and error bar represents the mean and SEM of the methylation grade difference.

Fig. 7 Schematic illustration of the CpG methylation status dislodged dependent on the extent of chromosomal losses. A methylation gradient between the undermethylated CpG-rich promoter region (CpG island) and the overmethylated CpG-poor region (non-island CpG) is prone to both hypo- and hypermethylation. The high-level (LOH-H) and low-level (LOH-L) chromosomal losses stimulate the variable hypomethylation in the methylation-gradient regions via dosage-compensation mechanism. Significant hypomethylation in the LOH-H cases demethylate the initial hypermethylation, leading to few hypermethylation. Meanwhile, the LOH-L cases are under the influence of low-grade hypomethylation and retain the initial methylation changes.
Cited by 3 articles
-
Chromosomal Losses are Associated with Hypomethylation of the Gene-Control Regions in the Stomach with a Low Number of Active Genes
Yu-Chae Jung, Seung-Jin Hong, Young-Ho Kim, Sung-Ja Kim, Seok-Jin Kang, Sang-Wook Choi, Mun-Gan Rhyu
J Korean Med Sci. 2008;23(6):1068-1089. doi: 10.3346/jkms.2008.23.6.1068.DNA Methylation and Expression Patterns of Key Tissue-specific Genes in Adult Stem Cells and Stomach Tissues
Seung-Jin Hong, Moo-Il Kang, Jung-Hwan Oh, Yu-Chae Jung, Young-Ho Kim, Sung-Ja Kim, Seung-Hye Choi, Eun-Joo Seo, Sang-Wook Choi, Mun-Gan Rhyu
J Korean Med Sci. 2009;24(5):918-929. doi: 10.3346/jkms.2009.24.5.918.DNA Methylation Patterns of Ulcer-Healing Genes Associated with the Normal Gastric Mucosa of Gastric Cancers
Seung-Jin Hong, Jung-Hwan Oh, Yu-Chae Jung, Young-Ho Kim, Sung-Ja Kim, Seok-Jin Kang, Eun-Joo Seo, Sang-Wook Choi, Moo-Il Kang, Mun-Gan Rhyu
J Korean Med Sci. 2010;25(3):405-417. doi: 10.3346/jkms.2010.25.3.405.
Reference
-
1. Balmain A, Gray J, Ponder B. The genetics and genomics of cancer. Nat Genet. 2003. 33:238–244.
Article2. Lasko D, Cavenee W, Nordenskjold M. Loss of constitutional heterozygosity in human cancer. Annu Rev Genet. 1991. 25:281–314.
Article3. Vogelstein B, Fearon ER, Kern SE, Hamilton SR, Preisinger AC, Nakamura Y, White R. Allelotype of colorectal carcinomas. Science. 1989. 244:207–211.
Article4. Choi SW, Lee KJ, Bae YA, Min KO, Kwon MS, Kim KM, Rhyu MG. Genetic classification of colorectal cancer based on chromosomal loss and microsatellite instability predicts survival. Clin Cancer Res. 2002. 8:2311–2322.5. Nagel S, Borisch B, Thein SL, Oestreicher M, Nothiger F, Birrer S, Tobler A, Fey MF. Somatic mutations detected by mini- and microsatellite DNA markers reveal clonal intratumor heterogeneity in gastrointestinal cancers. Cancer Res. 1995. 55:2866–2870.6. Chung YJ, Choi JR, Park SW, Kim KM, Rhyu MG. Evidence for two modes of allelic loss: multifocal analysis on both early and advanced gastric carcinomas. Virchows Arch. 2001. 438:31–38.
Article7. Kim KM, Kwon MS, Hong SJ, Min KO, Seo EJ, Lee KY, Choi SW, Rhyu MG. Genetic classification of intestinal-type and diffuse-type gastric cancers based on chromosomal loss and microsatellite instability. Virchows Arch. 2003. 443:491–500.
Article8. Hong SJ, Choi SW, Lee KH, Lee S, Min KO, Rhyu MG. Preoperative genetic diagnosis of gastric carcinoma based on chromosomal loss and microsatellite instability. Int J Cancer. 2005. 113:249–258.
Article9. Meller VH. Dosage compensation: making 1X equal 2X. Trends Cell Biol. 2000. 10:54–59.
Article10. Muller H. Why polyploid is rarer in animals than in plants. American Nature. 1925. 59:346–353.11. Gallardo MH, Bickham JW, Honeycutt RL, Ojeda RA, Kohler N. Discovery of tetraploidy in a mammal. Nature. 1999. 401:341.
Article12. Ehrlich M. DNA methylation in cancer: too much, but also too little. Oncogene. 2002. 21:5400–5413.
Article13. Lin CH, Hsieh SY, Sheen IS, Lee WC, Chen TC, Shyu WC, Liaw YF. Genome-wide hypomethylation in hepatocellular carcinogenesis. Cancer Res. 2001. 61:4238–4243.14. Henikoff S. Heterochromatin function in complex genomes. Biochim Biophys Acta. 2000. 1470:1–8.
Article15. Lee JH, Park SJ, Abraham SC, Seo JS, Nam JH, Choi C, Juhng SW, Rashid A, Hamilton SR, Wu TT. Frequent CpG island methylation in precursor lesions and early gastric adenocarcinomas. Oncogene. 2004. 23:4646–4654.
Article16. Ward RL, Cheong K, Ku SL, Meagher A, O'Connor T, Hawkins NJ. Adverse prognostic effect of methylation in colorectal cancer is reversed by microsatellite instability. J Clin Oncol. 2003. 21:3729–3736.
Article17. Yang AS, Estecio MR, Garcia-Manero G, Kantarjian HM, Issa JP. Comment on "Chromosomal instability and tumors promoted by DNA hypomethylation" and "Induction of tumors in mice by genomic hypomethylation". Science. 2003. 302:1153.
Article18. Lauren P. The two histological main types of gastric carcinoma: diffuse and so-called intestinal-type carcinoma. Acta Pathol Microbiol Scand. 1965. 64:31–49.19. Greene FL, Page DL, Fleming ID, Fritz A, Balch CM, Haller DG, Morrow M. AJCC cancer staging maunal. 2002. Berlin Heidelberg New York: Springer Verlag.20. Clark SJ, Harrison J, Paul CL, Frommer M. High sensitivity mapping of methylated cytosines. Nucleic Acids Res. 1994. 22:2990–2997.21. Herman JG, Umar A, Polyak K, Graff JR, Ahuja N, Issa JP, Markowitz S, Willson JK, Hamilton SR, Kinzler KW, Kane MF, Kolodner RD, Vogelstein B, Kunkel TA, Baylin SB. Incidence and functional consequences of hMLH1 promoter hypermethylation in colorectal carcinoma. Proc Natl Acad Sci USA. 1998. 95:6870–6875.
Article22. Lo KW, Kwong J, Hui AB, Chan SY, To KF, Chan AS, Chow LS, Teo PM, Johnson PJ, Huang DP. High frequency of promoter hypermethylation of RASSF1A in nasopharyngeal carcinoma. Cancer Res. 2001. 61:3877–3881.23. Herman JG, Graff JR, Myohanen S, Nelkin BD, Baylin SB. Methylation-specific PCR: a novel PCR assay for methylation status of CpG islands. Proc Natl Acad Sci USA. 1996. 93:9821–9826.
Article24. Kaneda A, Tsukamoto T, Takamura-Enya T, Watanabe N, Kaminishi M, Sugimura T, Tatematsu M, Ushijima T. Frequent hypomethylation in multiple promoter CpG islands is associated with global hypomethylation, but not with frequent promoter hypermethylation. Cancer Sci. 2004. 95:58–64.
Article25. Kokkola A, Monni O, Puolakkainen P, Nordling S, Haapiainen R, Kivilaakso E, Knuutila S. Presence of high-level DNA copy number gains in gastric carcinoma and severely dysplastic adenomas but not in moderately dysplastic adenomas. Cancer Genet Cytogenet. 1998. 107:32–36.
Article26. Akiyama Y, Maesawa C, Ogasawara S, Terashima M, Masuda T. Cell-type-specific repression of the maspin gene is disrupted frequently by demethylation at the promoter region in gastric intestinal metaplasia and cancer cells. Am J Pathol. 2003. 163:1911–1919.
Article27. Cho B, Lee H, Jeong S, Bang YJ, Lee HJ, Hwang KS, Kim HY, Lee YS, Kang GH, Jeoung DI. Promoter hypomethylation of a novel cancer/testis antigen gene CAGE is correlated with its aberrant expression and is seen in premalignant stage of gastric carcinoma. Biochem Biophys Res Commun. 2003. 307:52–63.
Article28. Martin L, Assem M, Piard F. Are there several types of colorectal carcinomas? Correlations with genetic data. Eur J Cancer Prev. 1999. 8:Suppl 1. 13–20.
Article29. Bariol C, Suter C, Cheong K, Ku SL, Meagher A, Hawkins N, Ward R. The relationship between hypomethylation and CpG island methylation in colorectal neoplasia. Am J Pathol. 2003. 162:1361–1371.
Article30. Kwon MS, Hong SJ, Cho HA, Ahn GH, Lee SS, Lee KY, Rhyu MG. Extensive and divergent chromosomal losses in squamous and spindle-cell components of esophageal sarcomatoid carcinoma. Virchows Arch. 2003. 443:635–642.
Article31. Kim KM, Kim MJ, Cho BK, Choi SW, Rhyu MG. Genetic evidence for the multi-step progression of mixed glandular-neuroendocrine gastric carcinomas. Virchows Arch. 2002. 440:85–93.
Article32. Velicescu M, Weisenberger DJ, Gonzales FA, Tsai YC, Nguyen CT, Jones PA. Cell division is required for de novo methylation of CpG islands in bladder cancer cells. Cancer Res. 2002. 62:2378–2384.33. Oue N, Oshimo Y, Nakayama H, Ito R, Yoshida K, Matsusaki K, Yasui W. DNA methylation of multiple genes in gastric carcinoma: association with histological type and CpG island methylator phenotype. Cancer Sci. 2003. 94:901–905.
Article34. Toyooka S, Toyooka KO, Harada K, Miyajima K, Makarla P, Sathyanarayana UG, Yin J, Sato F, Shivapurkar N, Meltzer SJ, Gazdar AF. Aberrant methylation of the CDH13 (H-cadherin) promoter region in colorectal cancers and adenomas. Cancer Res. 2002. 62:3382–3386.35. Song SH, Jong HS, Choi HH, Kang SH, Ryu MH, Kim NK, Kim WH, Bang YJ. Methylation of specific CpG sites in the promoter region could significantly down-regulate p16(INK4a) expression in gastric adenocarcinoma. Int J Cancer. 2000. 87:236–240.
Article36. Kang GH, Lee S, Shim YH, Kim JC, Ro JY. Profile of methylated CpG sites of hMLH1 promoter in primary gastric carcinoma with microsatellite instability. Pathol Int. 2002. 52:764–768.37. Sarbia M, Geddert H, Klump B, Kiel S, Iskender E, Gabbert HE. Hypermethylation of tumor suppressor genes (p16INK4A, p14ARF and APC) in adenocarcinomas of the upper gastrointestinal tract. Int J Cancer. 2004. 111:224–228.38. Lee TL, Leung WK, Chan MW, Ng EK, Tong JH, Lo KW, Chung SC, Sung JJ, To KF. Detection of gene promoter hypermethylation in the tumor and serum of patients with gastric carcinoma. Clin Cancer Res. 2002. 8:1761–1766.39. Waki T, Tamura G, Tsuchiya T, Sato K, Nishizuka S, Motoyama T. Promoter methylation status of E-cadherin, hMLH1, and p16 genes in nonneoplastic gastric epithelia. Am J Pathol. 2002. 161:399–403.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Chromosomal Losses are Associated with Hypomethylation of the Gene-Control Regions in the Stomach with a Low Number of Active Genes
- Methylation and Chromosomal Losses in Squamous Cell Carcinoma of the Head and Neck
- The Extent of Chromosomal Losses and the Status of CpG Methylation in Squamous Cell Carcinoma of the Head and Neck
- CpG Island Hypermethylation in Gastric Carcinoma and Its Premalignant Lesions
- DNA Methylation as Surrogate Marker For Gastric Cancer
