Potential Mechanism and Involvement of p120-Catenin in the Malignant Biology of Glioma
- Affiliations
-
- 1Department of Neurosurgery, Tianjin Medical University General Hospital, Tianjin, China
- 2Department of Neurosurgery, Cangzhou Central Hospital, Cangzhou, China
- 3Department of Emergency, Cangzhou Central Hospital, Cangzhou, China
- 4Department of Pathology, Cangzhou Central Hospital, Cangzhou, China
- KMID: 2560403
- DOI: http://doi.org/10.3340/jkns.2024.0053
Abstract
Objective
: This study analyzed the influence of p120-catenin (catenin [cadherin-associated protein], delta 1 [CTNND1]) on the malignant characteristics of glioma and elucidated the potential underlying mechanism.
Methods
: The p120 expression level was assessed in the brain tissues of 42 glioma patients and 10 patients with epilepsy by using the immunohistochemical method. Meanwhile, quantitative polymerase chain reaction (QT-PCR) technology was employed to assess the expression of p120 in the brain tissues of 71 glioma patients and 13 epilepsy patients. LN229, U251, and U87 glioma cells were used for in vitro analysis and categorized into four treatment groups : siRNA-blank control (BC) group (no RNA sequence was transfected), siRNA-negative control (NC) group (transfected control RNA sequences with no effect), and siRNA-1 and siRNA-2 groups (two p120-specific interfering RNA transfection). p120 expression in these treatment groups was quantified by western blotting assay. The migratory and invasive capabilities of glioma cells were studied by wound healing assay and Transwell invasion assay, respectively, under different treatment conditions. MTT (3-(4,5)-dimethylthiahiazo (-z-y1)-3,5-di-phenytetrazoliumromide) assay and cell cycle and apoptosis assay were used to determine glioma cell proliferation and apoptosis, respectively. Enzymelabeled assay was performed to measure intracellular calcium ion concentration. Immunofluorescence assay was performed for determining microtubule formation and glioma cell distribution.
Results
: Brain tissues of the glioma group exhibited a remarkable increase in the p120 expression level as compared to brain tissues of the nontumor group (p<0.05). Furthermore, a strong positive correlation was noted between the malignancy degree in glioma brain tissues and p120 expression in Western blotting (r=0.906, p<0.0001) and QT-PCR (F=830.6, p<0.01). Compared to the BC and NC groups, the siRNA transfection groups showed a significant suppression in p120 expression in glioma cells (p<0.05), with a marked attenuation in the invasive, migratory, and proliferative capabilities of glioma cells as well as an increase in apoptotic potential (p<0.05). Enzyme-labeled assay showed a remarkable increase in calcium concentration in glioma cells after siRNA treatment. Immunofluorescence assay revealed that the microtubule formation ability of glioma cells reduced after siRNA treatment.
Conclusion
: p120 has a pivotal involvement in facilitating glioma cell invasion and proliferation by potentially modulating these processes through its involvement in microtubule formation and regulation of intracellular calcium ion levels.
Keyword
Figure
-
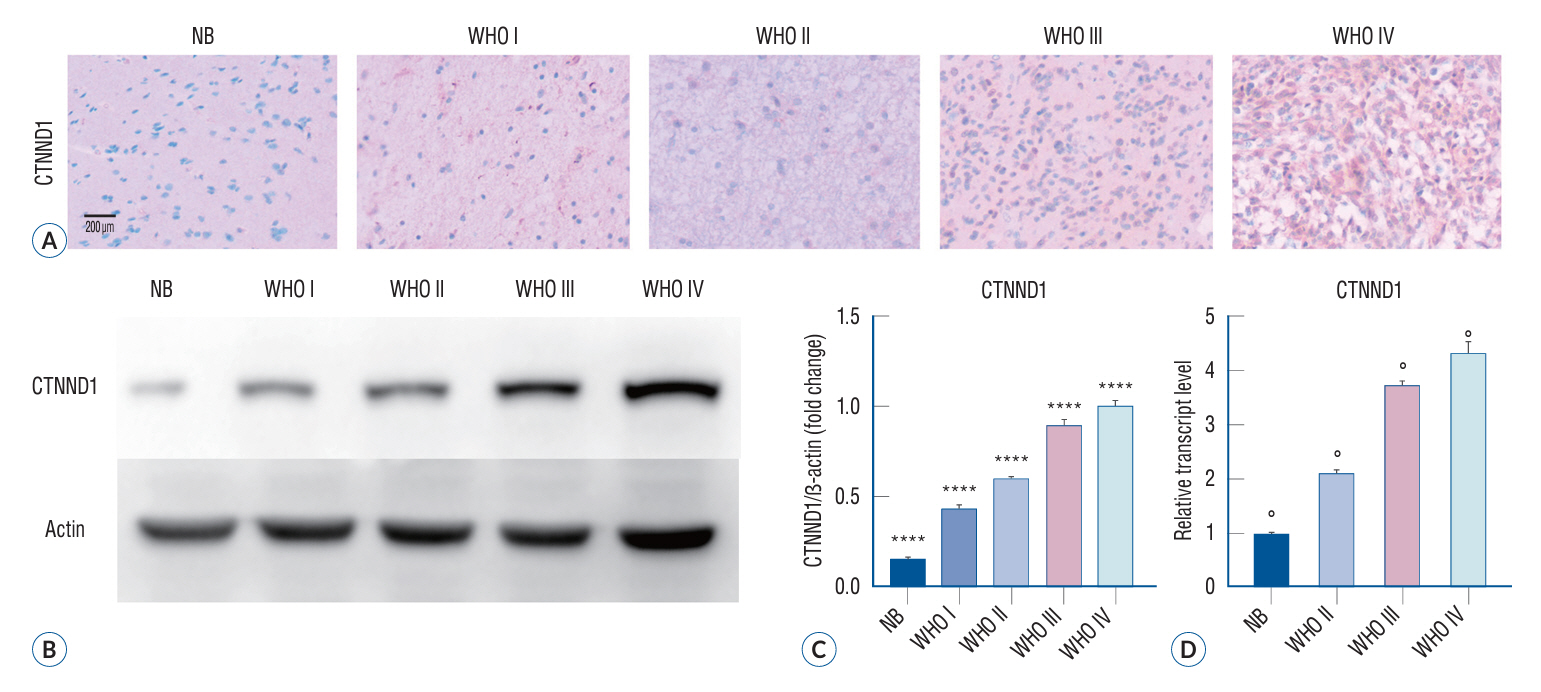
Fig. 1. Expression of p120 in human glioma specimens. A : The cytoplasmic localization of CTNND1, as indicated by the presence of brown staining observed and photographed under a microscope, exhibited a positive association with the malignancy level of glioma. B : Western blot analysis was employed to assess the presence of the two proteins in healthy brain tissues and various stages of glioma tissues (p<0.05). C : The Pearson correlation test was conducted to analyze the association between CTNND1 expression and the malignancy level of glioma specimens. The findings revealed a positive correlation between CTNND1 expression and the degree of malignancy in glioma (r=0.906; ****p<0.0001). D : The results of quantitative PCR indicated a high expression of CTNND1 in gliomas, which was positively associated with the grade of glioma (F=830.6, °p<0.01). NB : normal brain tissue, WHO : World Health Organization, CTNND1 : catenin (cadherin-associated protein), delta 1.
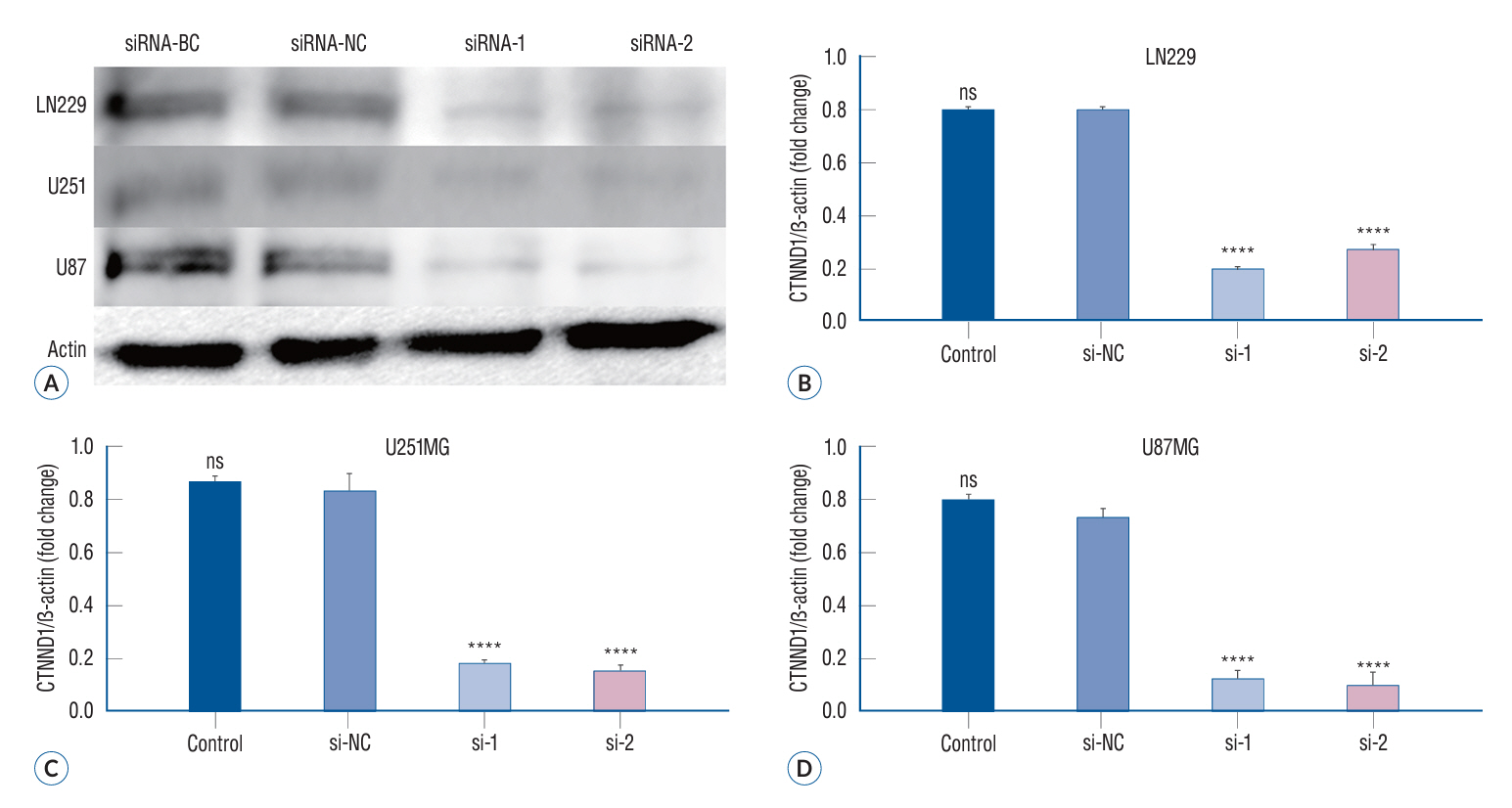
Fig. 2. The expression of p120 in different treatment groups of glioma cells. With GAPDH as internal reference, the expression of p120 in siRNA-1 and siRNA-2 groups was significantly lower than that in siRNA-BC and siRNA-NC groups (A-D) (****p<0.0001). BC : blank control group, NC : negative control group, CTNND1 : catenin (cadherin-associated protein), delta 1, ns : no significance, GAPDH : glyceraldehyde-3-phosphate dehydrogenase.
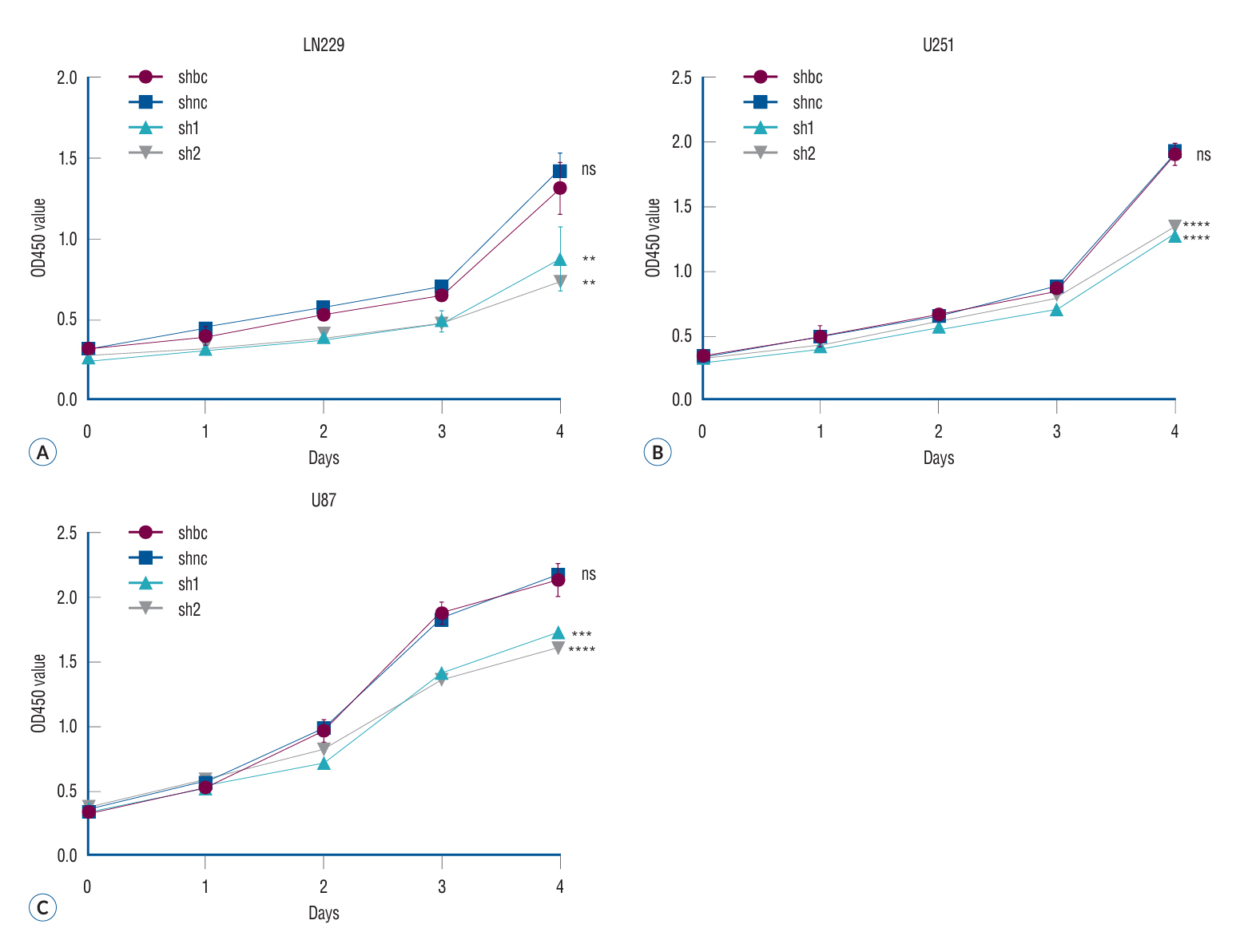
Fig. 3. The proliferation of glioma cells in different treatment groups was evaluated by MTT assay. The MTT method, also referred to as MTT colorimetry, is a technique utilized for assessing cell viability and proliferation. The underlying detection principle involves the reduction of exogenous MTT by succinate dehydrogenase in the mitochondria of viable cells, resulting in the formation and deposition of water-insoluble blue-purple crystal Formazan within the cells. Conversely, non-viable cells lack this functionality. Dimethyl sulfoxide can dissolve the Formazan present in the cells, and subsequently, an enzyme-linked immunosorbent assay determines the light absorption value at either 540 or 720 nm wavelengths. The results showed that the proliferation ability of the two glioma cells decreased significantly after siRNA in the three malignant glioma cell lines (A-C). **p<0.01. ***p<0.001. ****p<0.001. shbc : siRNA blank controlshnc, shnc : siRNA normal control, sh1 : siRNA-1 group, sh2 : siRNA-2 group, ns : no significance, MTT : 3-(4,5)-dimethylthiahiazo (-z-y1)-3,5-diphenytetrazoliumromide.
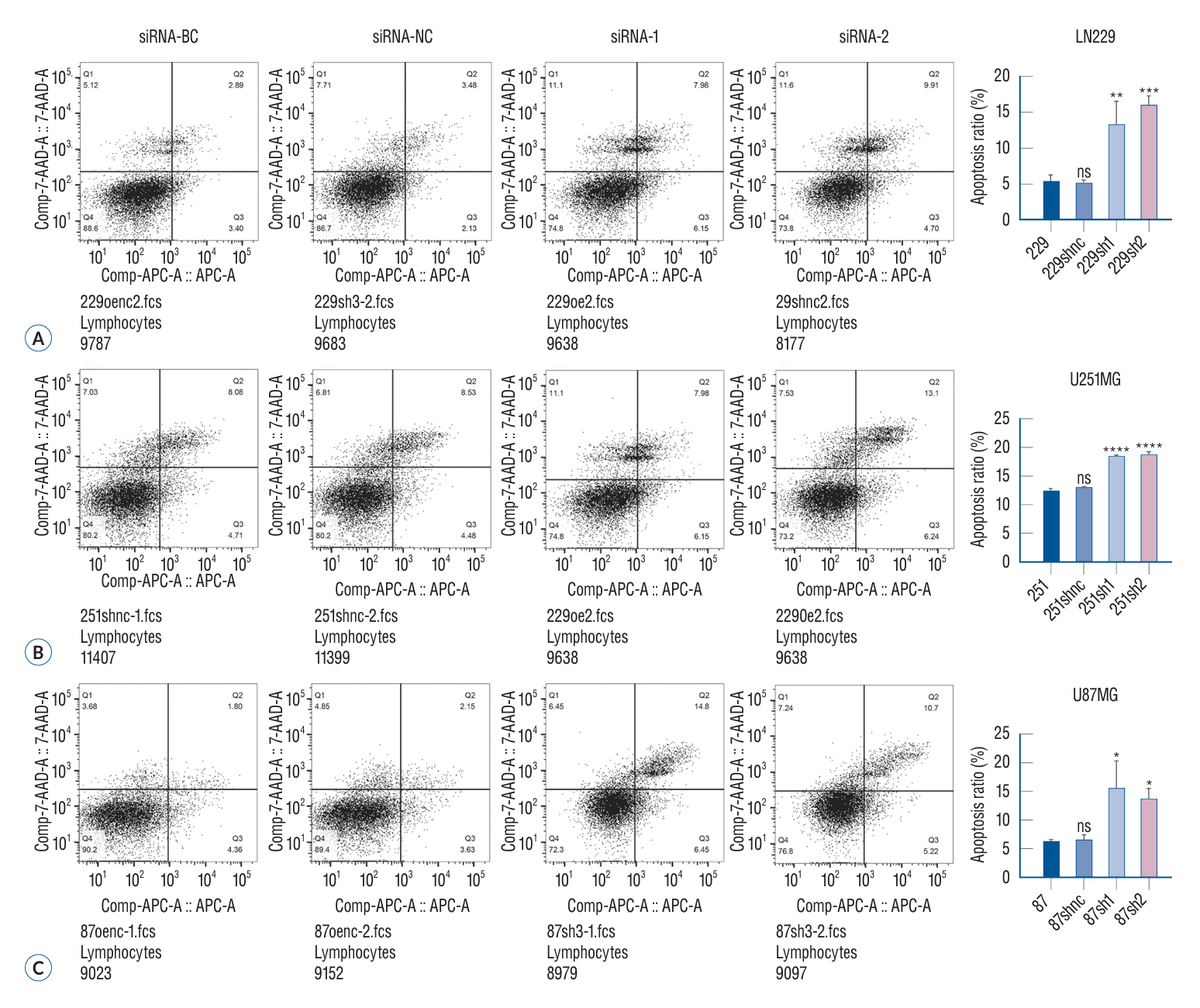
Fig. 4. The apoptosis of glioma cells in different treatment groups was detected by flow cytometry. Flow cytometry is a common method to measure apoptosis. The principle is that when apoptosis occurs, the permeability of the cell membrane also increases, but the degree is between normal cells and necrotic cells. Using this feature, the examined cell suspension was stained with fluorescein, and the fluorescence intensity in the cell suspension was measured by flow cytometry to distinguish normal cells from necrotic cells and apoptotic cells. The results showed that the apoptosis ability of the two glioma cells increased significantly after siRNA treatment (A-C). *p<0.05. **p<0.01. ***p<0.001. ****p<0.001. BC : blank control group, NC : negative control group, 7-AADA-A : 7-aminoactinomycin D apoptosis, APC-A : allophycocyanin apoptosis, ns : no significance.

Fig. 5. The migration ability of different treatment groups of glioma cells evaluated using a wound healing assay. After a 24-hour period of treatment involving the application of specific siRNA, a wound was created using the end of a sterile pipette with a volume capacity of 200 μL. Following this, the culture medium was substituted with serum-free medium and the cells were subsequently cultured for an additional 24 hours before being captured in photographs. A : In comparison to the siRNA-BC and siRNA-NC groups, the wound size was observed to be greater in the siRNA-1 and siRNA-2 groups 24 hours after the formation of wounds. B-D : In vitro experiments demonstrated a significant reduction in glioma cell migration upon treatment with P120-specific siRNA, as indicated by quantitative data (****p<0.0001). BC : blank control group, NC : negative control group, ns : no significance.
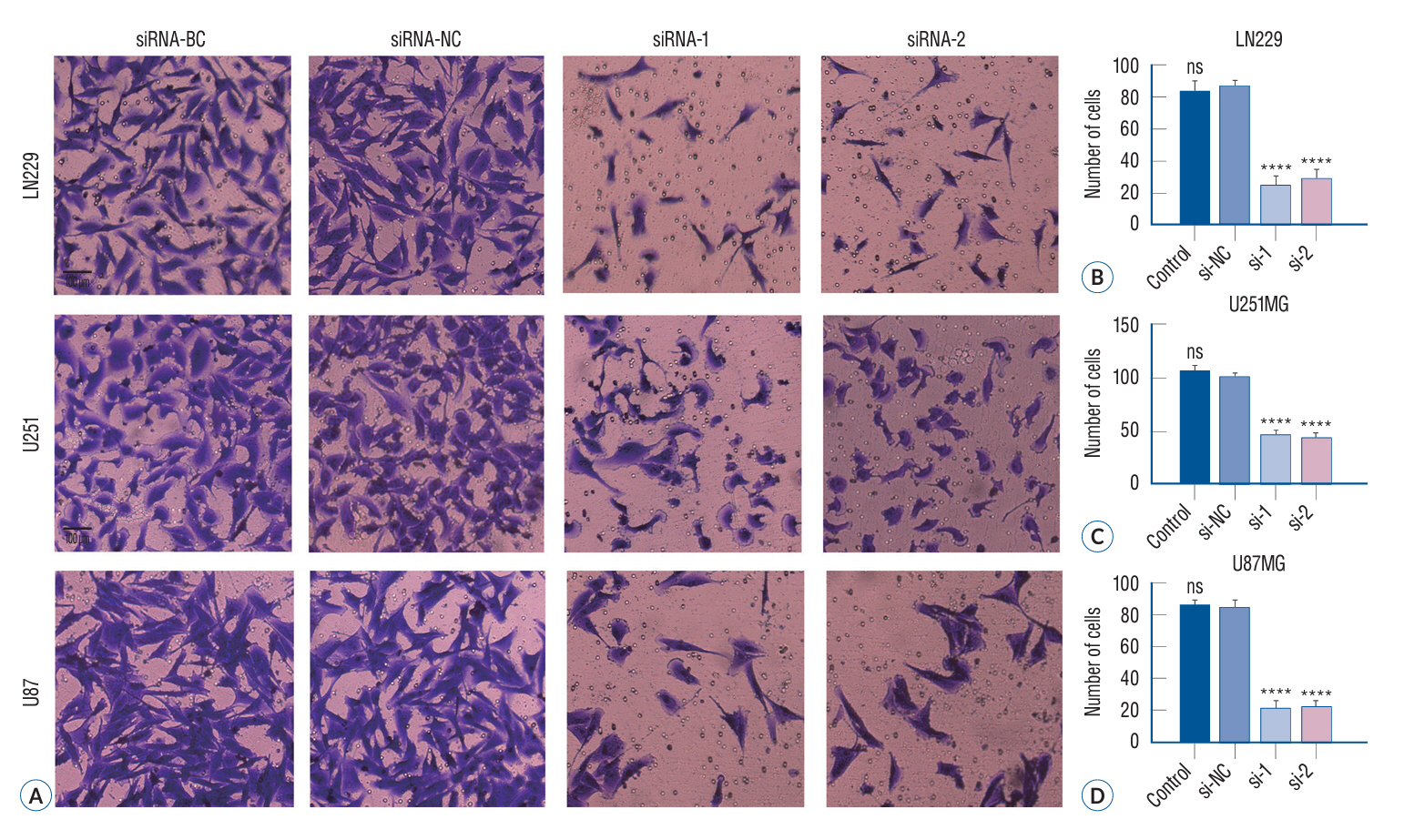
Fig. 6. Knock down of cortactin inhibited glioma cell invasion in vitro. Following a 24-hour treatment period, we conducted a Transwell invasion assay and subsequently incubated the cells for an additional 48-hour. The stroma-migrating cells were then stained with crystal violet and photographed. Representative images of glioma cells in the Millipore Matrigel-coated invasion chambers (A). Relevant quantitative data demonstrated a significant reduction in the invasion of glioma cells in vitro upon specific siRNA treatment (B-D) (****p<0.0001). BC : blank control group, NC : negative control group, ns : no significance.
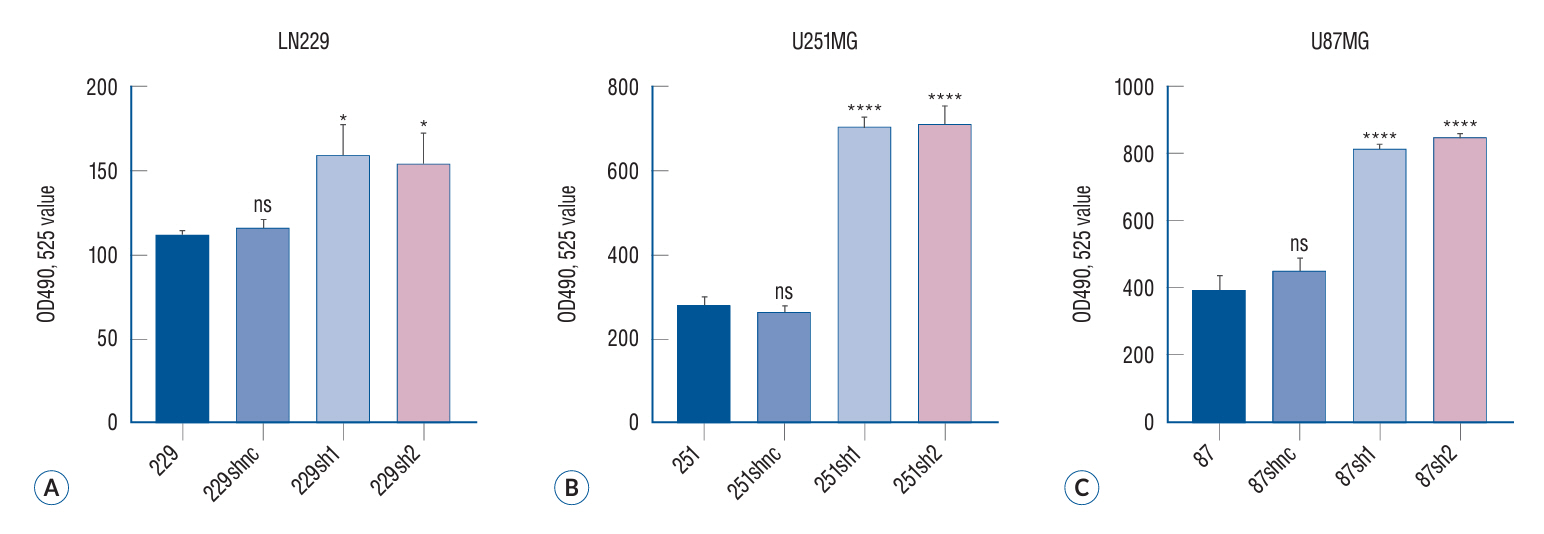
Fig. 7. The concentration of calcium ion in glioma cells of different treatment groups was detected by enzyme-labeled method. Calcium channel is the channel through which cells transport calcium ions. Calcium overload caused by the increase of intracellular calcium ion concentration can cause the disorder of oxidative phosphorylation in mitochondria, the decrease of mitochondrial membrane potential, the decrease of adenosine-triphosphate content in tissues, and the activation of phospholipase and protease in cytoplasm, which can lead to and promote irreversible damage of cells. After the deletion of p120 expression in glioma cells, the intracellular calcium concentration in both groups was significantly increased (A-C). *p<0.05. ****p<0.0001.
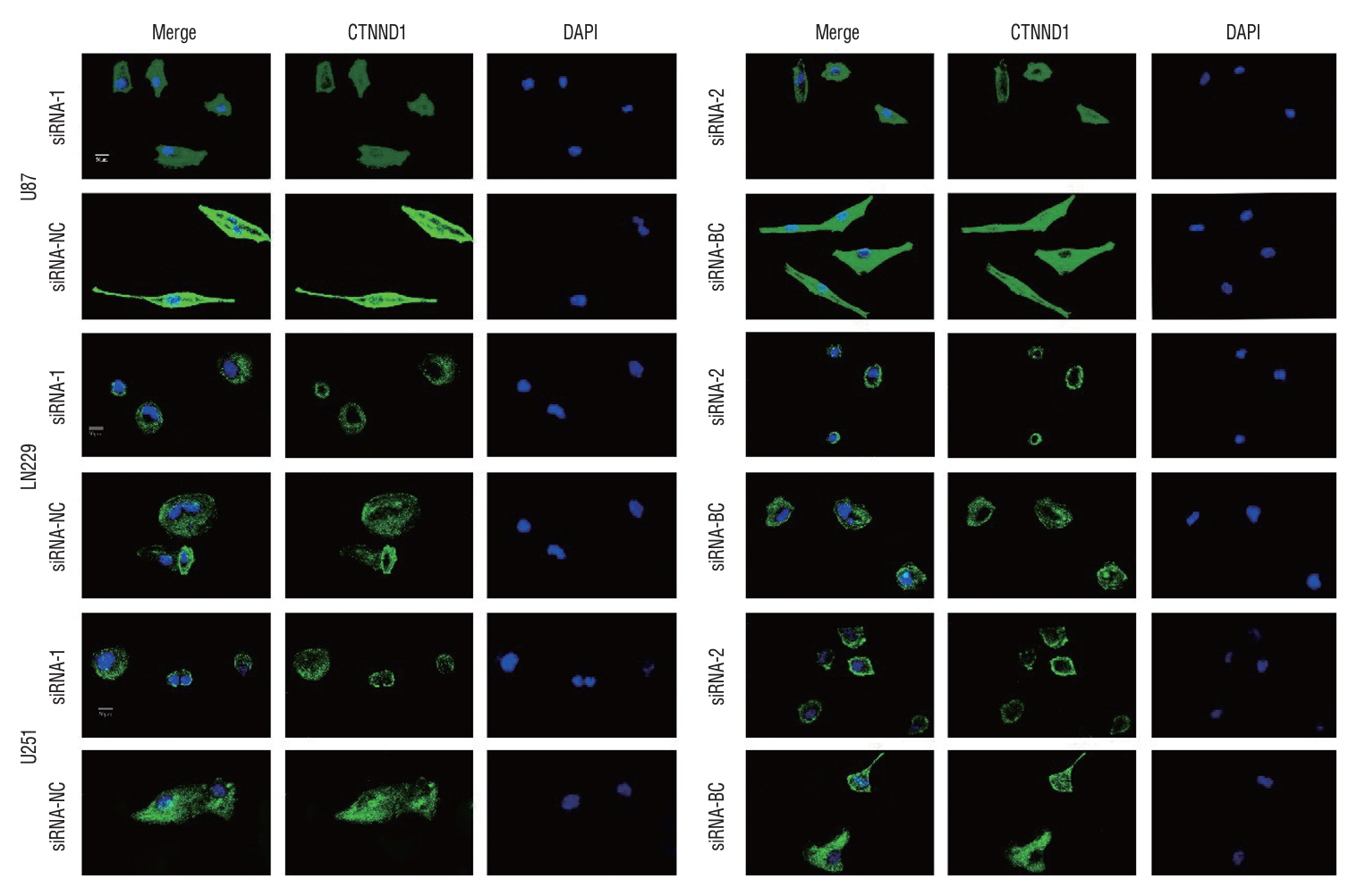
Fig. 8. The microtubule formation of glioma cells in different treatment groups was detected by immunofluorescence. To assess the inhibitory impact of siRNA targeting p120 on microtubule assembly in glioma cells, we conducted immunofluorescence staining for tubulin and DAPI to examine the nuclei of different treatment groups. The findings revealed a significant reduction in the ability of glioma cells to form microtubules following downregulation of p120 expression. These outcomes imply that p120 plays a crucial role in facilitating microtubule formation within glioma cells. The maglification is ×400. CTNND1 : catenin (cadherin-associated protein), delta 1, DAPI : 4ʹ,6-diamidino-2-phenylindole, NC : negative control group, BC : blank control group.
Cited by 1 articles
-
Editors’ Pick in November 2024
Bum-Tae Kim
J Korean Neurosurg Soc. 2024;67(6):593-594. doi: 10.3340/jkns.2024.0190.
Reference
-
References
1. Barthel L, Hadamitzky M, Dammann P, Schedlowski M, Sure U, Thakur BK, et al. Glioma: molecular signature and crossroads with tumor microenvironment. Cancer Metastasis Rev. 41:53–75. 2022.
Article2. Becchetti A. Chapter two - interplay of Ca2+ and K+ signals in cell physiology and cancer. Curr Top Membr. 92:15–46. 2023.3. Boiarska Z, Passarella D. Microtubule-targeting agents and neurodegeneration. Drug Discov Today. 26:604–615. 2021.4. Di Cristofori A, Basso G, de Laurentis C, Mauri I, Sirtori MA, Ferrarese C, et al. Perspectives on (a)symmetry of arcuate fasciculus. A short review about anatomy, tractography and TMS for arcuate fasciculus reconstruction in planning surgery for gliomas in language areas. Front Neurol. 12:639822. 2021.5. Duñach M, Del Valle-Pérez B, García de Herreros A. p120-catenin in canonical Wnt signaling. Crit Rev Biochem Mol Biol. 52:327–339. 2017.
Article6. Goodson HV, Jonasson EM. Microtubules and microtubule-associated proteins. Cold Spring Harb Perspect Biol. 10:a022608. 2018.
Article7. Haag D, Zipper P, Westrich V, Karra D, Pfleger K, Toedt G, et al. Nos2 inactivation promotes the development of medulloblastoma in Ptch1(+/-) mice by deregulation of Gap43-dependent granule cell precursor migration. PLoS Genet. 8:e1002572. 2012.
Article8. Hatanaka K, Simons M, Murakami M. Phosphorylation of VE-cadherin controls endothelial phenotypes via p120-catenin coupling and Rac1 activation. Am J Physiol Heart Circ Physiol. 300:H162–H172. 2011.
Article9. Hernández A, Domènech M, Muñoz-Mármol AM, Carrato C, Balana C. Glioblastoma: relationship between metabolism and immunosuppressive microenvironment. Cells. 10:3529. 2021.
Article10. Hong JY, Oh IH, McCrea PD. Phosphorylation and isoform use in p120-catenin during development and tumorigenesis. Biochim Biophys Acta. 1863:102–114. 2016.
Article11. Jiang G, Wang Y, Dai S, Liu Y, Stoecker M, Wang E, et al. P120-catenin isoforms 1 and 3 regulate proliferation and cell cycle of lung cancer cells via β-catenin and Kaiso respectively. PLoS One. 7:e30303. 2012.
Article12. Jiang Y, Liao L, Shrestha C, Ji S, Chen Y, Peng J, et al. Reduced expression of E-cadherin and p120-catenin and elevated expression of PLC-γ1 and PIKE are associated with aggressiveness of oral squamous cell carcinoma. Int J Clin Exp Pathol. 8:9042–9051. 2015.13. Jung E, Osswald M, Ratliff M, Dogan H, Xie R, Weil S, et al. Tumor cell plasticity, heterogeneity, and resistance in crucial microenvironmental niches in glioma. Nat Commun. 12:1014. 2021.
Article14. Krigers A, Demetz M, Moser P, Kerschbaumer J, Brawanski KR, Fritsch H, et al. Impact of GAP-43, Cx43 and actin expression on the outcome and overall survival in diffuse and anaplastic gliomas. Sci Rep. 13:2024. 2023.
Article15. Latario CJ, Schoenfeld LW, Howarth CL, Pickrell LE, Begum F, Fischer DA, et al. Tumor microtubes connect pancreatic cancer cells in an Arp2/3 complex-dependent manner. Mol Biol Cell. 31:1259–1272. 2020.
Article16. Lee M, Ji H, Furuta Y, Park JI, McCrea PD. p120-catenin regulates REST and CoREST, and modulates mouse embryonic stem cell differentiation. J Cell Sci. 127(Pt 18):4037–4051. 2014.17. Lou E. A ticket to ride: the implications of direct intercellular communication via tunneling nanotubes in peritoneal and other invasive malignancies. Front Oncol. 10:559548. 2020.
Article18. Maiden SL, Petrova YI, Gumbiner BM. Microtubules inhibit E-cadherin adhesive activity by maintaining phosphorylated p120-catenin in a colon carcinoma cell model. PLoS One. 11:e0148574. 2016.
Article19. Osswald M, Jung E, Sahm F, Solecki G, Venkataramani V, Blaes J, et al. Brain tumour cells interconnect to a functional and resistant network. Nature. 528:93–98. 2015.
Article20. Osswald M, Solecki G, Wick W, Winkler F. A malignant cellular network in gliomas: potential clinical implications. Neuro Oncol. 18:479–485. 2016.
Article21. Parato J, Bartolini F. The microtubule cytoskeleton at the synapse. Neurosci Lett. 753:135850. 2021.
Article22. Poff A, Koutnik AP, Egan KM, Sahebjam S, D’Agostino D, Kumar NB. Targeting the Warburg effect for cancer treatment: ketogenic diets for management of glioma. Semin Cancer Biol. 56:135–148. 2019.
Article23. Portela M, Mitchell T, Casas-Tintó S. Cell-to-cell communication mediates glioblastoma progression in Drosophila. Biol Open. 9:bio053405. 2020.24. Przybylowski CJ, Hervey-Jumper SL, Sanai N. Surgical strategy for insular glioma. J Neurooncol. 51:491–497. 2021.
Article25. Roehlecke C, Schmidt MHH. Tunneling nanotubes and tumor microtubes in cancer. Cancers (Basel). 12:857. 2020.
Article26. Saberbaghi T, Wong R, Rutka JT, Wang GL, Feng ZP, Sun HS. Role of Cl-channels in primary brain tumour. Cell Calcium. 81:1–11. 2019.27. Venhuizen JH, Jacobs FJC, Span PN, Zegers MM. P120 and E-cadherin: double-edged swords in tumor metastasis. Semin Cancer Biol. 60:107–120. 2020.
Article28. Venkataramani V, Schneider M, Giordano FA, Kuner T, Wick W, Herrlinger U, et al. Disconnecting multicellular networks in brain tumours. Nat Rev Cancer. 22:481–491. 2022.
Article29. Venkataramani V, Yang Y, Schubert MC, Reyhan E, Tetzlaff SK, Wißmann N, et al. Glioblastoma hijacks neuronal mechanisms for brain invasion. Cell. 185:2899–2917.e31. 2022.
Article30. Venkatesh VS, Lou E. Tunneling nanotubes: a bridge for heterogeneity in glioblastoma and a new therapeutic target? Cancer Rep (Hoboken). 2:e1185. 2019.
Article31. Wang X, Liang J, Sun H. The network of tumor microtubes: an improperly reactivated neural cell network with stemness feature for resistance and recurrence in gliomas. Front Oncol. 12:921975. 2022.
Article32. Watson DC, Bayik D, Storevik S, Moreino SS, Sprowls SA, Han J, et al. GAP43-dependent mitochondria transfer from astrocytes enhances glioblastoma tumorigenicity. Nat Cancer. 4:648–664. 2023.
Article33. Weil S, Osswald M, Solecki G, Grosch J, Jung E, Lemke D, et al. Tumor microtubes convey resistance to surgical lesions and chemotherapy in gliomas. Neuro Oncol. 19:1316–1326. 2017.
Article34. Wu H, Yan X, Zhao L, Li X, Li X, Zhang Y, et al. p120-catenin promotes innate antiviral immunity through stabilizing TBK1-IRF3 complex. Mol Immunol. 157:8–17. 2023.
Article35. Xie R, Kessler T, Grosch J, Hai L, Venkataramani V, Huang L, et al. Tumor cell network integration in glioma represents a stemness feature. Neuro Oncol. 23:757–769. 2021.
Article36. Xie Z, Tang Y, Man MQ, Shrestha C, Bikle DD. p120-catenin is required for regulating epidermal proliferation, differentiation, and barrier function. J Cell Physiol. 234:427–432. 2018.
Article37. Xu S, Tang L, Li X, Fan F, Liu Z. Immunotherapy for glioma: current management and future application. Cancer Lett. 476:1–12. 2020.
Article38. Yasinjan F, Xing Y, Geng H, Guo R, Yang L, Liu Z, et al. Immunotherapy: a promising approach for glioma treatment. Front Immunol. 14:1255611. 2023.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Current Immunotherapeutic Approaches for Malignant Gliomas
- Inhibition of Wnt/ββ-catenin signaling by monensin in cervical cancer
- A Case of Brain Stem Glioma Responded to Tamoxifen after Relapse with Chemotherapy and Radiotherapy
- Myosin VI contributes to malignant proliferation of human glioma cells
- Impact of General Factors on Glioma Immunotherapy
