Glucagon-Like Peptide Receptor Agonist Inhibits Angiotensin II-Induced Proliferation and Migration in Vascular Smooth Muscle Cells and Ameliorates Phosphate-Induced Vascular Smooth Muscle Cells Calcification
- Affiliations
-
- 1Institute of Medical Research, Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine, Seoul, Korea
- 2Division of Endocrinology and Metabolism, Department of Internal Medicine, Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine, Seoul, Korea
- KMID: 2551265
- DOI: http://doi.org/10.4093/dmj.2022.0363
Abstract
- Background
Glucagon-like peptide-1 receptor agonist (GLP-1RA), which is a therapeutic agent for the treatment of type 2 diabetes mellitus, has a beneficial effect on the cardiovascular system.
Methods
To examine the protective effects of GLP-1RAs on proliferation and migration of vascular smooth muscle cells (VSMCs), A-10 cells exposed to angiotensin II (Ang II) were treated with either exendin-4, liraglutide, or dulaglutide. To examine the effects of GLP-1RAs on vascular calcification, cells exposed to high concentration of inorganic phosphate (Pi) were treated with exendin-4, liraglutide, or dulaglutide.
Results
Ang II increased proliferation and migration of VSMCs, gene expression levels of Ang II receptors AT1 and AT2, proliferation marker of proliferation Ki-67 (Mki-67), proliferating cell nuclear antigen (Pcna), and cyclin D1 (Ccnd1), and the protein expression levels of phospho-extracellular signal-regulated kinase (p-Erk), phospho-c-JUN N-terminal kinase (p-JNK), and phospho-phosphatidylinositol 3-kinase (p-Pi3k). Exendin-4, liraglutide, and dulaglutide significantly decreased the proliferation and migration of VSMCs, the gene expression levels of Pcna, and the protein expression levels of p-Erk and p-JNK in the Ang II-treated VSMCs. Erk inhibitor PD98059 and JNK inhibitor SP600125 decreased the protein expression levels of Pcna and Ccnd1 and proliferation of VSMCs. Inhibition of GLP-1R by siRNA reversed the reduction of the protein expression levels of p-Erk and p-JNK by exendin-4, liraglutide, and dulaglutide in the Ang II-treated VSMCs. Moreover, GLP-1 (9-36) amide also decreased the proliferation and migration of the Ang II-treated VSMCs. In addition, these GLP-1RAs decreased calcium deposition by inhibiting activating transcription factor 4 (Atf4) in Pi-treated VSMCs.
Conclusion
These data show that GLP-1RAs ameliorate aberrant proliferation and migration in VSMCs through both GLP-1Rdependent and independent pathways and inhibit Pi-induced vascular calcification.
Keyword
Figure
-
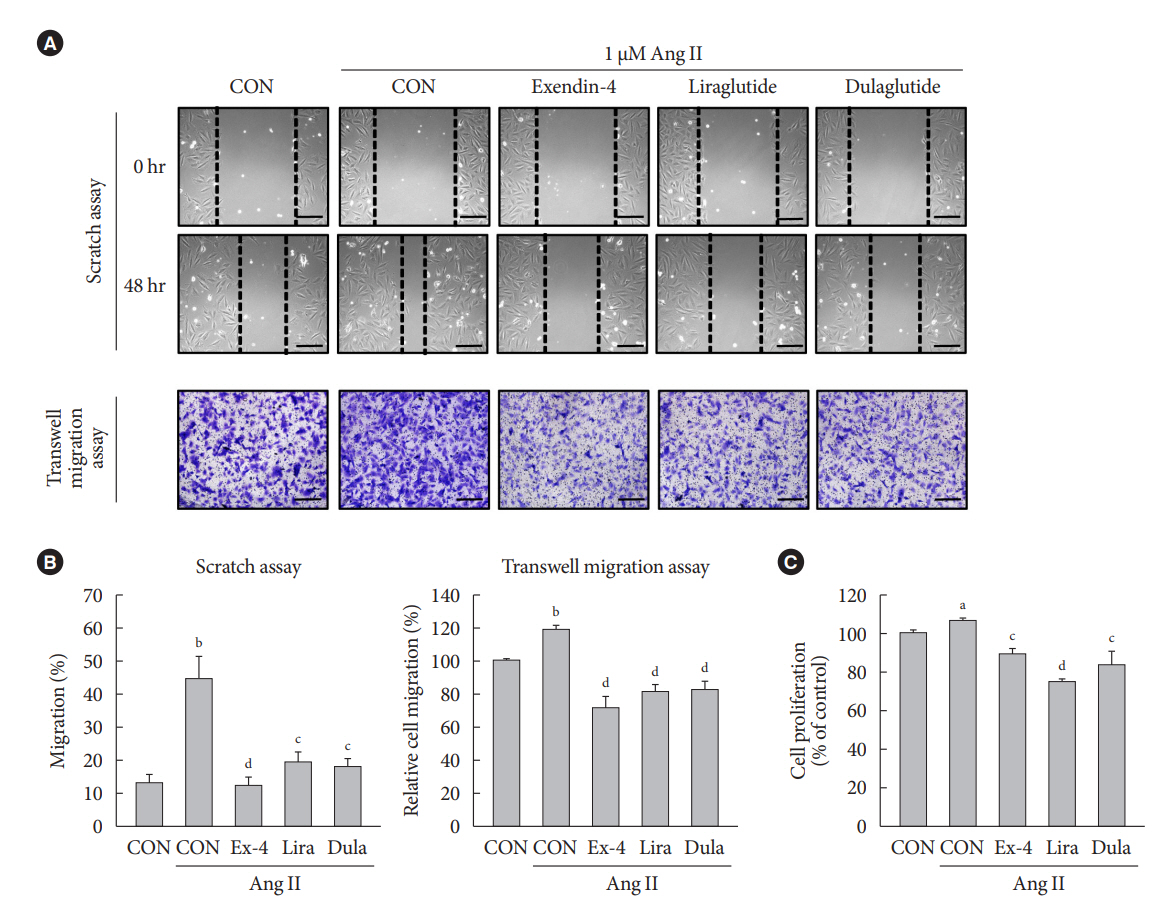
Fig. 1. Exendin-4, liraglutide, and dulaglutide inhibit the migration and proliferation of vascular smooth muscle cells (VSMCs) treated with angiotensin II (Ang II). A-10 cells are treated with 1 μM Ang II, followed by treatment with or without 100 nM exendin-4 (Ex-4), liraglutide (Lira), and dulaglutide (Dula) for 48 hours. (A, B) VSMC migration is determined using scratch wound healing assay and transwell migration assay, and (C) VSMC proliferation is determined using MTT assay. aP<0.05 and bP<0.01 when compared with the control cells, cP<0.05 and dP<0.01 when compared with the Ang II-treated cells.
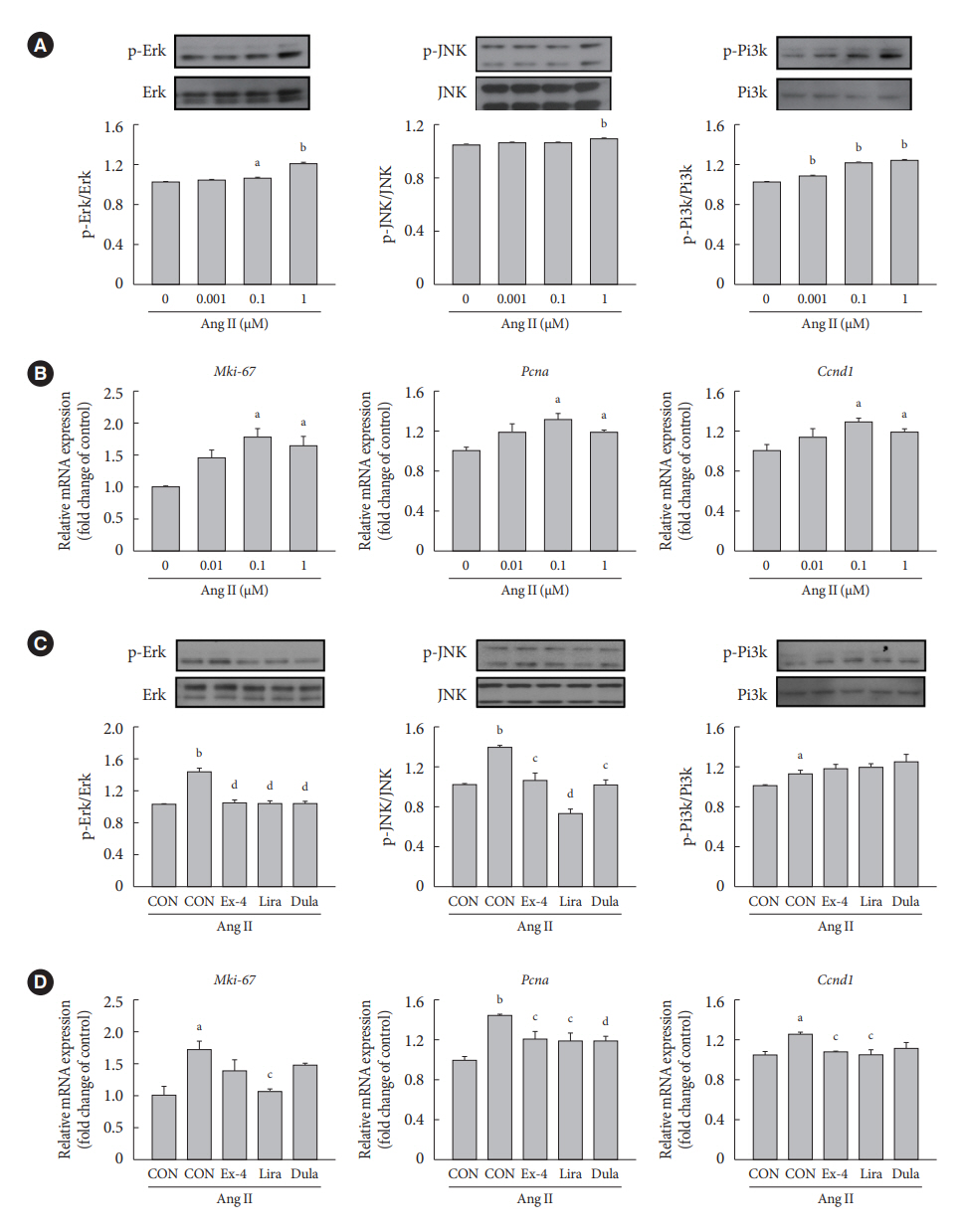
Fig. 2. Exendin-4, liraglutide, and dulaglutide inhibit extracellular signal-regulated kinase (Erk) and c-JUN N-terminal kinase (JNK) signaling pathways and the expression of proliferation marker genes in vascular smooth muscle cells (VSMCs) treated with angiotensin II (Ang II). (A, B) A-10 cells are treated with various concentrations of Ang II for 24 hours. (C, D) A-10 cells are treated with 1 μM Ang II, followed by treatment with or without 100 nM exendin-4 (Ex-4), liraglutide (Lira), and dulaglutide (Dula) for 24 hours. p-Erk, p-JNK, and phospho-phosphatidylinositol 3-kinase (p-Pi3k) levels are analyzed using Western blotting. The mRNA expression levels of the genes encoding marker of proliferation Ki-67 (Mki-67), proliferating cell nuclear antigen (Pcna), and cyclin D1 (Ccnd1) are analyzed with quantitative real-time polymerase chain reaction and normalized to that of the glyceraldehyde3-phosphate dehydrogenase (Gapdh) gene. aP<0.05 and bP<0.01 when compared with the control (CON) cells, cP<0.05 and dP<0.01 when compared with the Ang II-treated cells.
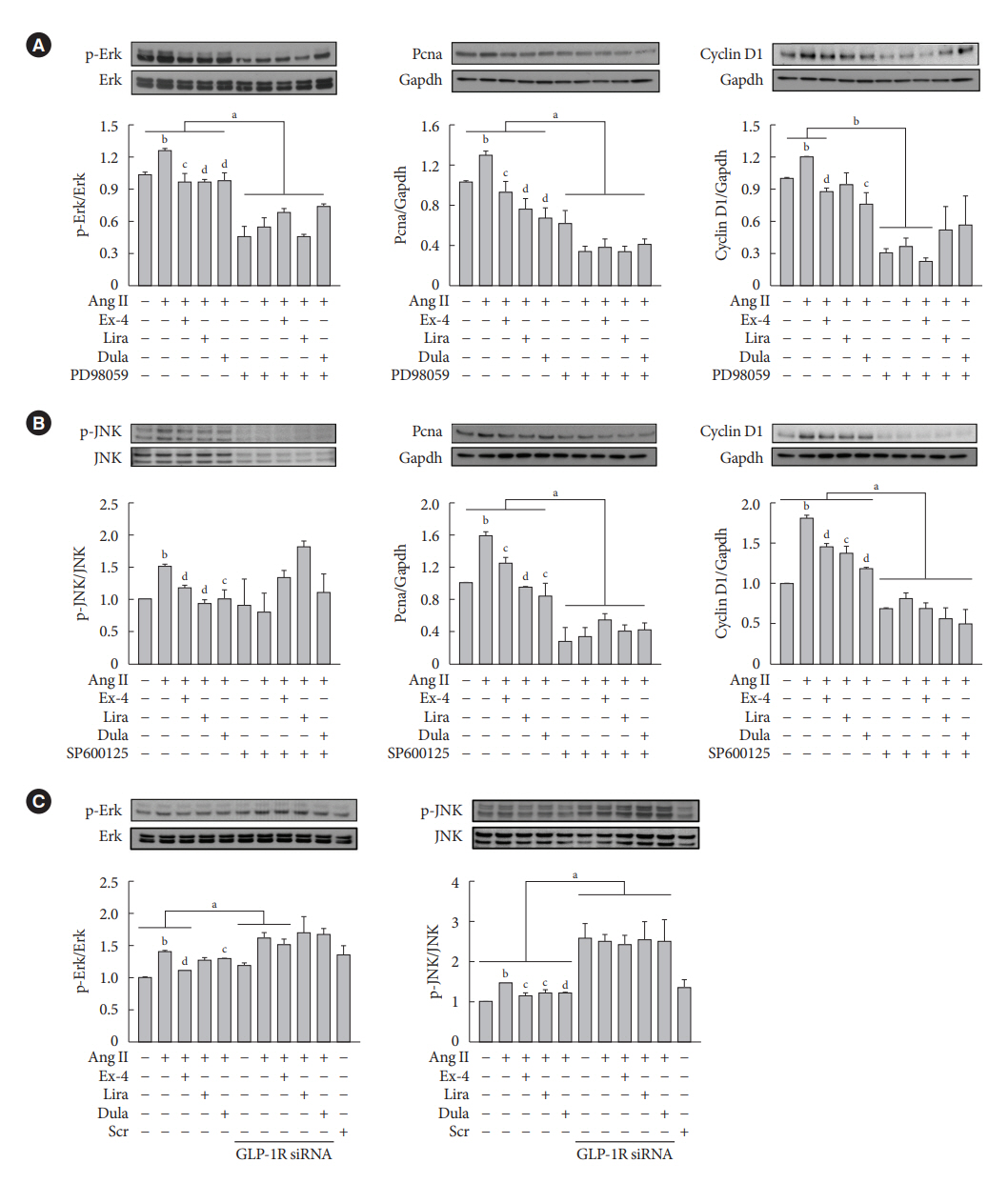
Fig. 3. Inhibitory effects of exendin-4 (Ex-4), liraglutide (Lira), and dulaglutide (Dula) on the expression of extracellular signal-regulated kinase (Erk) and c-JUN N-terminal kinase (JNK) are mediated by glucagon-like peptide-1 receptor (GLP-1R). A-10 cells pre-exposed to (A) 50 μM Erk inhibitor (PD98059) or (B) 50 μM JNK inhibitor (SP600125) for 1 hour are treated with 1 μM angiotensin II (Ang II), followed by treatment with or without Ex-4 (100 nM), Lira (100 nM), and Dula (100 nM) for 24 hours. (C) A-10 cells, transfected with 50 nM GLP-1R siRNA or scrambled (Scr) siRNA for 24 hours, are treated with Ang II, followed by treatment with or without Ex-4, Lira, and Dula for 24 hours. Expression levels of phosphor-Erk (p-Erk), p-JNK, proliferating cell nuclear antigen (Pcna), and cyclin D1 are analyzed using Western blotting, and glyceraldehyde-3-phosphate dehydrogenase (Gapdh) is used as the loading control. aP<0.05 and bP<0.01 when compared with the control cells, cP<0.05 and dP<0.01 when compared with the Ang II-treated cells.
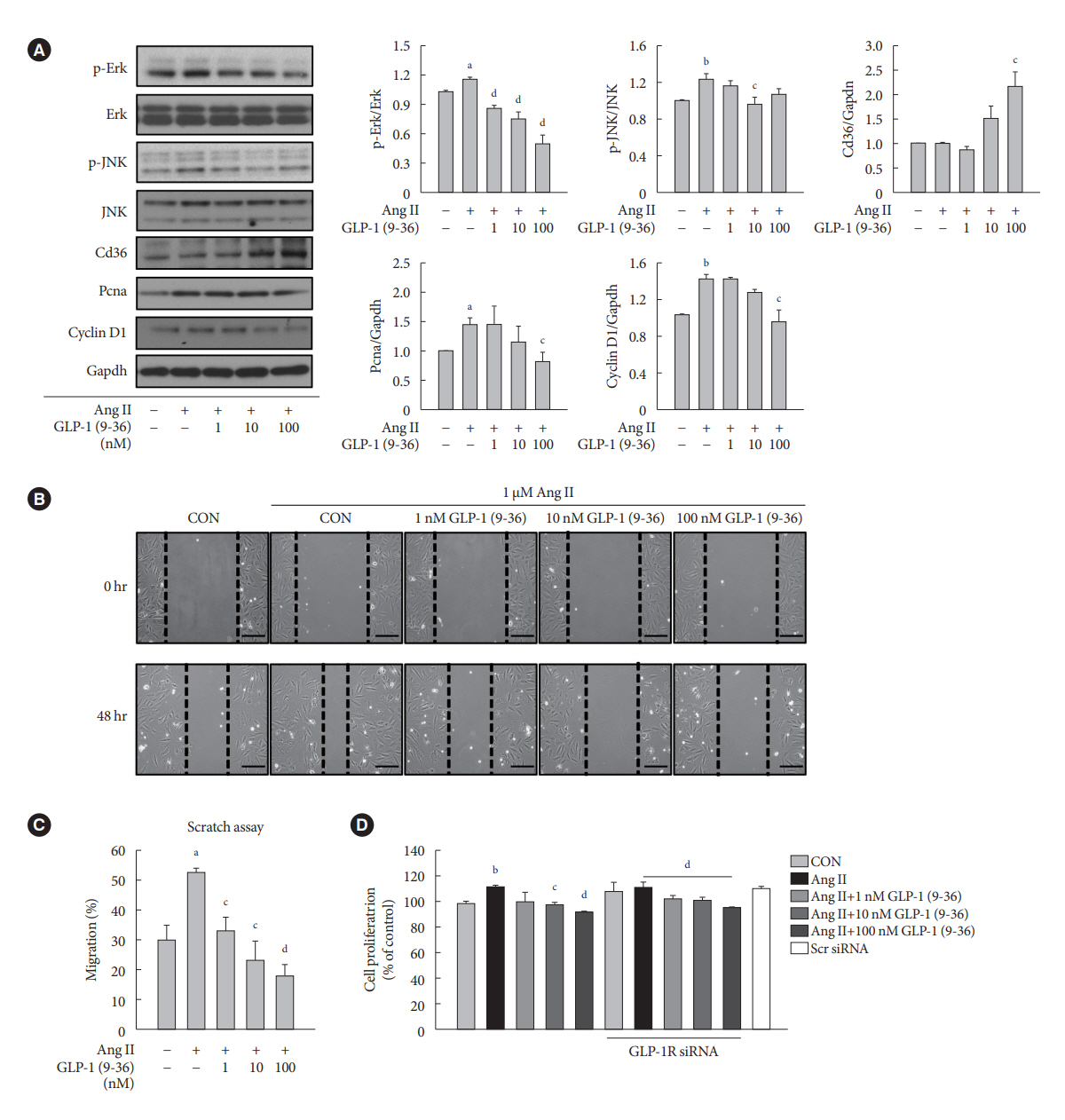
Fig. 4. Glucagon-like peptide-1 (GLP-1) (9-36) amide decreases the proliferation and migration of vascular smooth muscle cells (VSMCs) treated with angiotensin II (Ang II) by increasing Cd36 expression. A-10 cells are treated with 1 μM Ang II, followed by treatment with various concentrations of GLP-1 (9-36) amide (1 to 100 nM) for 24 or 48 hours. (A) Expression levels of phosphoextracellular signal-regulated kinase (p-Erk), phospho-c-JUN N-terminal kinase (p-JNK), Cd36, proliferating cell nuclear antigen (Pcna), and cyclin D1 are analyzed using Western blotting, and glyceraldehyde-3-phosphate dehydrogenase (Gapdh) is used as the loading control (CON). (B, C) VSMC migration is determined using scratch wound healing assay and quantified. (D) A-10 cells transfected with 50 nM GLP-1 receptor (GLP-1R) siRNA or scramble (Scr) siRNA are treated with 1 μM Ang II, followed by treatment with various concentrations of GLP-1 (9-36) amide (1 to 100 nM) for 24 hours. VSMC proliferation is determined using MTT assay. aP<0.05 and bP<0.01 when compared with the CON cells, cP<0.05 and dP<0.01 when compared with the Ang II-treated cells.
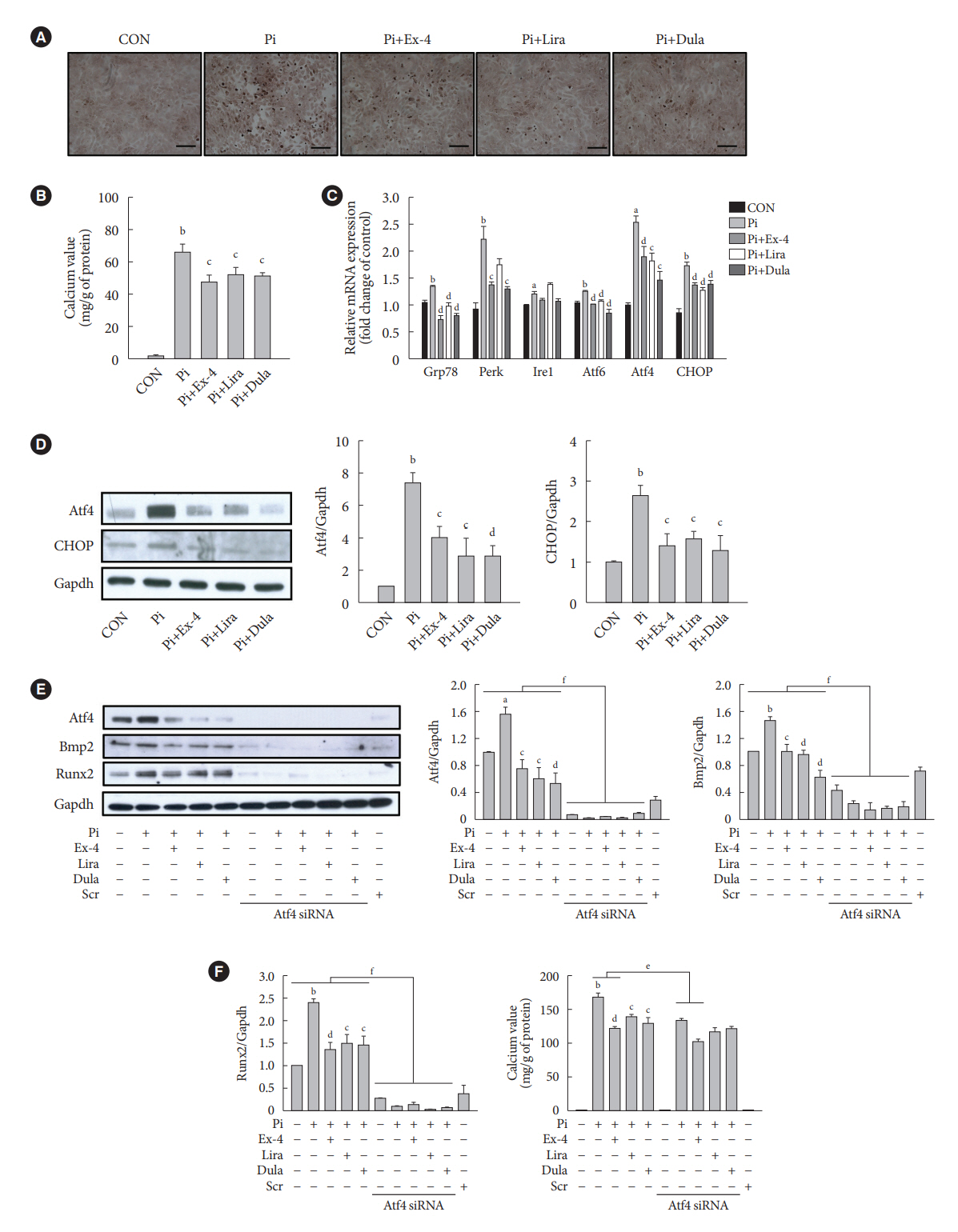
Fig. 5. Exendin-4 (Ex-4), liraglutide (Lira), and dulaglutide (Dula) inhibit high inorganic phosphate (Pi)-induced vascular calcification in vascular smooth muscle cells. A-10 cells are treated with 4 mM Pi, followed by treatment with or without 100 nM Ex-4, Lira, or Dula for 7 days. Levels of calcium deposition are assessed by (A) alizarin red S staining and (B) calcium assays. (C, D) The protein and gene expressions of endoplasmic reticulum (ER) stress markers are analyzed using Western blotting and quantitative real-time polymerase chain reaction, respectively. (E, F) A-10 cells transfected with 10 nM activating transcription factor 4 (Atf4) siRNA or scramble (Scr) siRNA are treated with 4 mM Pi, followed by treatment with or without 100 nM Ex-4, Lira, or Dula. Expression levels of Atf4, bone morphogenic protein 2 (Bmp2), and runt-related transcription factor-2 (Runx2) are analyzed using Western blotting, and glyceraldehyde-3-phosphate dehydrogenase (Gapdh) is used as the loading control (CON). Levels of calcium deposition are assessed by calcium assays. Grp78, 78 kDa glucose-regulated protein; Perk, protein kinase RNA-like endoplasmic reticulum kinase; Ire1, inositol-requiring protein 1; CHOP, C/EBP homologous protein. aP<0.05 and bP<0.01 when compared with the CON cells, cP<0.05 and dP<0.01 when compared with the Pi-treated cells, eP<0.05 and fP<0.01 when compared with each siRNA-untreated groups.
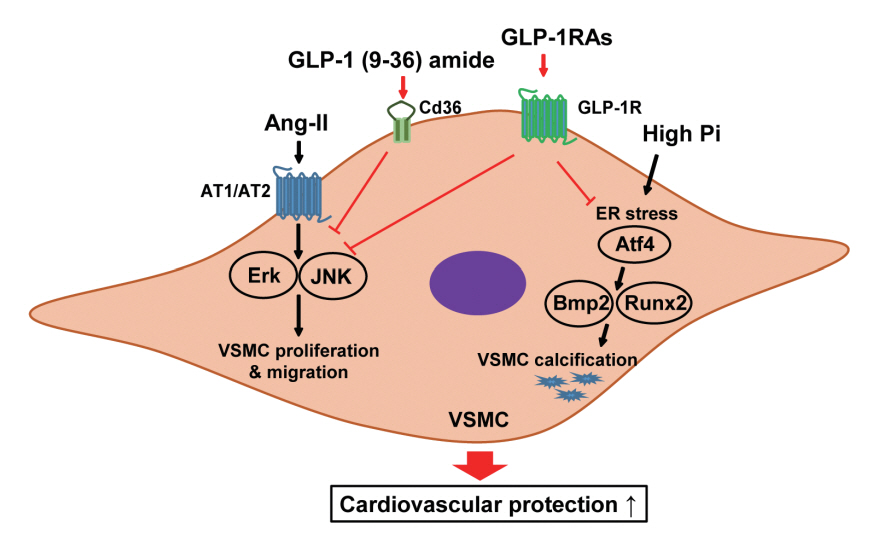
Fig. 6. Schematic diagram illustrating a possible mechanism of the protective effect of glucagon-like peptide-1 receptor agonists (GLP-1RAs) on cardiovascular disease. Ang II, angiotensin II; Pi, inorganic phosphate; AT1, angiotensin II receptor type 1; AT2, angiotensin II receptor type 2; Erk, extracellular signal-regulated kinase; JNK, c-JUN N-terminal kinase; VSMC, vascular smooth muscle cell; ER, endoplasmic reticulum; Atf4, activating transcription factor 4; Bmp2, bone morphogenic protein 2; Runx2, runtrelated transcription factor-2.
Reference
-
1. Anderson SL, Marrs JC. Antihyperglycemic medications and cardiovascular risk reduction. Eur Endocrinol. 2017; 13:86–90.2. Pozo L, Bello F, Suarez A, Ochoa-Martinez FE, Mendez Y, Chang CH, et al. Novel pharmacological therapy in type 2 diabetes mellitus with established cardiovascular disease: current evidence. World J Diabetes. 2019; 10:291–303.3. Naito R, Miyauchi K. Coronary artery disease and type 2 diabetes mellitus. Int Heart J. 2017; 58:475–80.4. Basatemur GL, Jorgensen HF, Clarke MCH, Bennett MR, Mallat Z. Vascular smooth muscle cells in atherosclerosis. Nat Rev Cardiol. 2019; 16:727–44.5. van Diepen JA, Berbee JF, Havekes LM, Rensen PC. Interactions between inflammation and lipid metabolism: relevance for efficacy of anti-inflammatory drugs in the treatment of atherosclerosis. Atherosclerosis. 2013; 228:306–15.6. Wang Y, Dubland JA, Allahverdian S, Asonye E, Sahin B, Jaw JE, et al. Smooth muscle cells contribute the majority of foam cells in ApoE (apolipoprotein E)-deficient mouse atherosclerosis. Arterioscler Thromb Vasc Biol. 2019; 39:876–87.7. Grootaert MOJ, Bennett MR. Vascular smooth muscle cells in atherosclerosis: time for a re-assessment. Cardiovasc Res. 2021; 117:2326–39.8. Bennett MR, Sinha S, Owens GK. Vascular smooth muscle cells in atherosclerosis. Circ Res. 2016; 118:692–702.9. van Thiel BS, van der Pluijm I, te Riet L, Essers J, Danser AH. The renin-angiotensin system and its involvement in vascular disease. Eur J Pharmacol. 2015; 763(Pt A):3–14.10. Dasgupta C, Zhang L. Angiotensin II receptors and drug discovery in cardiovascular disease. Drug Discov Today. 2011; 16:22–34.11. Lemarie CA, Schiffrin EL. The angiotensin II type 2 receptor in cardiovascular disease. J Renin Angiotensin Aldosterone Syst. 2010; 11:19–31.12. Yang J, Chen C, Ren H, Han Y, He D, Zhou L, et al. Angiotensin II AT(2) receptor decreases AT(1) receptor expression and function via nitric oxide/cGMP/Sp1 in renal proximal tubule cells from Wistar-Kyoto rats. J Hypertens. 2012; 30:1176–84.13. Guimond MO, Gallo-Payet N. How does angiotensin AT(2) receptor activation help neuronal differentiation and improve neuronal pathological situations? Front Endocrinol (Lausanne). 2012; 3:164.14. Chai W, Wang W, Dong Z, Cao W, Liu Z. Angiotensin II receptors modulate muscle microvascular and metabolic responses to insulin in vivo. Diabetes. 2011; 60:2939–46.15. Zhao Y, Shang F, Shi W, Zhang J, Zhang J, Liu X, et al. Angiotensin II receptor type 1 antagonists modulate vascular smooth muscle cell proliferation and migration via AMPK/mTOR. Cardiology. 2019; 143:1–10.16. Trion A, van der Laarse A. Vascular smooth muscle cells and calcification in atherosclerosis. Am Heart J. 2004; 147:808–14.17. Shi X, Gao J, Lv Q, Cai H, Wang F, Ye R, et al. Calcification in atherosclerotic plaque vulnerability: friend or foe? Front Physiol. 2020; 11:56.18. Proudfoot D, Shanahan CM. Biology of calcification in vascular cells: intima versus media. Herz. 2001; 26:245–51.19. Lanzer P, Boehm M, Sorribas V, Thiriet M, Janzen J, Zeller T, et al. Medial vascular calcification revisited: review and perspectives. Eur Heart J. 2014; 35:1515–25.20. Lau WL, Festing MH, Giachelli CM. Phosphate and vascular calcification: emerging role of the sodium-dependent phosphate co-transporter PiT-1. Thromb Haemost. 2010; 104:464–70.21. Saraiva FK, Sposito AC. Cardiovascular effects of glucagon-like peptide 1 (GLP-1) receptor agonists. Cardiovasc Diabetol. 2014; 13:142.22. Marso SP, Daniels GH, Brown-Frandsen K, Kristensen P, Mann JF, Nauck MA, et al. Liraglutide and cardiovascular outcomes in type 2 diabetes. N Engl J Med. 2016; 375:311–22.23. Marso SP, Bain SC, Consoli A, Eliaschewitz FG, Jodar E, Leiter LA, et al. Semaglutide and cardiovascular outcomes in patients with type 2 diabetes. N Engl J Med. 2016; 375:1834–44.24. Kieffer TJ, McIntosh CH, Pederson RA. Degradation of glucose-dependent insulinotropic polypeptide and truncated glucagon-like peptide 1 in vitro and in vivo by dipeptidyl peptidase IV. Endocrinology. 1995; 136:3585–96.25. Meier JJ, Gethmann A, Nauck MA, Gotze O, Schmitz F, Deacon CF, et al. The glucagon-like peptide-1 metabolite GLP-1-(9-36) amide reduces postprandial glycemia independently of gastric emptying and insulin secretion in humans. Am J Physiol Endocrinol Metab. 2006; 290:E1118–23.26. Li J, Zheng J, Wang S, Lau HK, Fathi A, Wang Q. Cardiovascular benefits of native GLP-1 and its metabolites: an indicator for GLP-1-therapy strategies. Front Physiol. 2017; 8:15.27. Ban K, Noyan-Ashraf MH, Hoefer J, Bolz SS, Drucker DJ, Husain M. Cardioprotective and vasodilatory actions of glucagonlike peptide 1 receptor are mediated through both glucagonlike peptide 1 receptor-dependent and -independent pathways. Circulation. 2008; 117:2340–50.28. Shi L, Ji Y, Jiang X, Zhou L, Xu Y, Li Y, et al. Liraglutide attenuates high glucose-induced abnormal cell migration, proliferation, and apoptosis of vascular smooth muscle cells by activating the GLP-1 receptor, and inhibiting ERK1/2 and PI3K/Akt signaling pathways. Cardiovasc Diabetol. 2015; 14:18.29. Sheahan KH, Wahlberg EA, Gilbert MP. An overview of GLP1 agonists and recent cardiovascular outcomes trials. Postgrad Med J. 2020; 96:156–61.30. Giugliano D, Scappaticcio L, Longo M, Caruso P, Maiorino MI, Bellastella G, et al. GLP-1 receptor agonists and cardiorenal outcomes in type 2 diabetes: an updated meta-analysis of eight CVOTs. Cardiovasc Diabetol. 2021; 20:189.31. Rhee EJ. Extra-glycemic effects of anti-diabetic medications: two birds with one stone? Endocrinol Metab (Seoul). 2022; 37:415–29.32. Caussy C, Aubin A, Loomba R. The relationship between type 2 diabetes, NAFLD, and cardiovascular risk. Curr Diab Rep. 2021; 21:15.33. Knudsen LB, Pridal L. Glucagon-like peptide-1-(9-36) amide is a major metabolite of glucagon-like peptide-1-(7-36) amide after in vivo administration to dogs, and it acts as an antagonist on the pancreatic receptor. Eur J Pharmacol. 1996; 318:429–35.34. Sonne DP, Engstrom T, Treiman M. Protective effects of GLP-1 analogues exendin-4 and GLP-1(9-36) amide against ischemia-reperfusion injury in rat heart. Regul Pept. 2008; 146:243–9.35. Tomas E, Habener JF. Insulin-like actions of glucagon-like peptide-1: a dual receptor hypothesis. Trends Endocrinol Metab. 2010; 21:59–67.36. Shao W, Wang Z, Ip W, Chiang YT, Xiong X, Chai T, et al. GLP1(28-36) improves β-cell mass and glucose disposal in streptozotocin-induced diabetic mice and activates cAMP/PKA/ β-catenin signaling in β-cells in vitro. Am J Physiol Endocrinol Metab. 2013; 304:E1263–72.37. Chistiakov DA, Sobenin IA, Orekhov AN, Bobryshev YV. Mechanisms of medial arterial calcification in diabetes. Curr Pharm Des. 2014; 20:5870–83.38. Lee J, Hong SW, Kim MJ, Kwon H, Park SE, Rhee EJ, et al. Metformin, resveratrol, and exendin-4 inhibit high phosphate-induced vascular calcification via AMPK-RANKL signaling. Biochem Biophys Res Commun. 2020; 530:374–80.39. Al-Aly Z. Phosphate, oxidative stress, and nuclear factor-κB activation in vascular calcification. Kidney Int. 2011; 79:1044–7.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Downregulation of Angiotensin II-Induced 12-Lipoxygenase Expression and Cell Proliferation in Vascular Smooth Muscle Cells from Spontaneously Hypertensive Rats by CCL5
- Losartan Inhibits Vascular Smooth Muscle Cell Proliferation through Activation of AMP-Activated Protein Kinase
- Role of zinc for calcification inhibitor protein in vascular smooth muscle cell plaque formation
- Monotropein resists atherosclerosis by reducing inflammation, oxidative stress, and abnormal proliferation and migration of vascular smooth muscle cells
- Pitavastatin Regulates Ang II Induced Proliferation and Migration via IGFBP-5 in VSMC
