A Hypoxia-Induced SCFFBXL1 E3 Ligase Ubiquitinates and Degrades the MEN1 Tumor Suppressor to Promote Colorectal Cancer Tumorigenesis
- Affiliations
-
- 1Department of Gastroenterology, Jiangxi Provincial People's Hospital Affiliated to Nanchang University, Nanchang, Jiangxi, China
- 2Department of Oncology, Jiangxi Provincial People's Hospital Affiliated to Nanchang University, Nanchang, Jiangxi, China
- KMID: 2528222
- DOI: http://doi.org/10.4143/crt.2021.373
Abstract
- Purpose
Emerging evidence has shown that SKP1-cullin-1-F-box-protein (SCF) E3 ligases contribute to the pathogenesis of different cancers by mediating the ubiquitination and degradation of tumor suppressors. However, the functions of SCF E3 ligases in the pathogenesis of colorectal cancer (CRC) remain obscure.
Materials and Methods
The cancerous and adjacent noncancerous tissues from CRC patients were collected, and protein levels were analyzed. Lentiviral short hairpin RNA (shRNA) and plasmid transfection were used to knock down and overexpress gene expression in CRC cell lines. Immunoprecipitation (IP), mass spectrometry, and co-IP analyses were used to determine protein interactions and the assembly of the SCF complex. Cell proliferation, migration, and tumor xenograft assays were performed to examine the effects of SCF members on CRC cell growth in vitro and in vivo.
Results
Hypoxia activated the docking of hypoxia-inducible factor 1α (HIF1α) onto the CUL1 promoter and induced CUL1 expression in CRC cells. CUL1 coupled with RBX1, SKP1, and FBXL1 to assemble the SCFFBXL1 complex in CRC biopsies and cells. The SCFFBXL1 E3 ligase specifically ubiquitinated and degraded the MEN1 tumor suppressor. Knockdown of HIF1α or SCFFBXL1 members, or blockage of SCFFBXL1 by two inhibitors (DT204 and SZLP1-41) caused the accumulation of MEN1 protein and led to a significant decrease in cell proliferation and migration in vitro and tumor growth in vivo.
Conclusion
The SCFFBXL1 E3 ligase is required for the ubiquitination of MEN1, and disruption of this complex may represent a new therapeutic strategy for the treatment of CRC.
Keyword
Figure
-
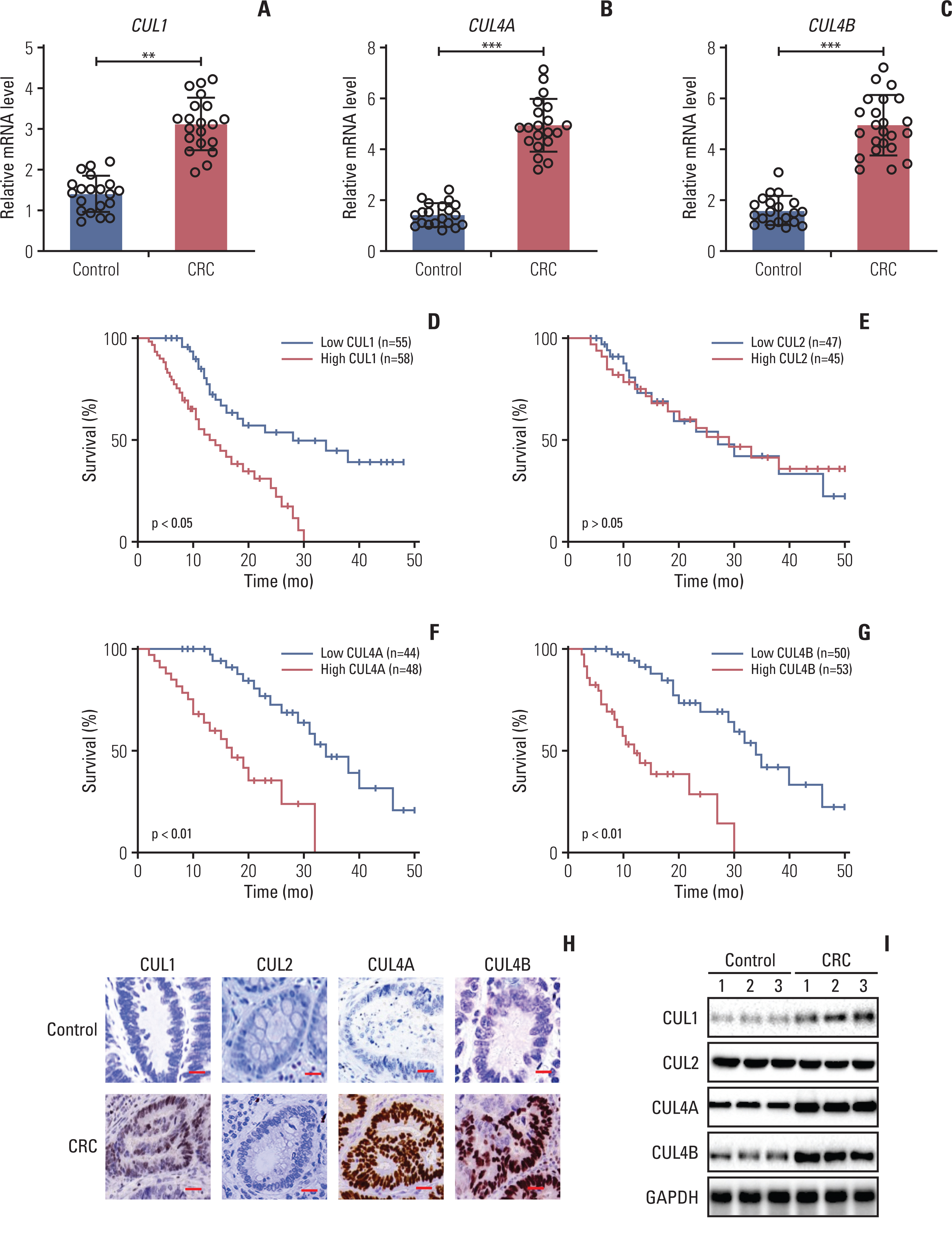
Fig. 1 CUL1 acted as an oncogene in colorectal cancer (CRC) tumor tissues. (A–C) The mRNA levels of CUL1 (A), CUL4A (B), and CUL4B (C) in tumor tissues. Twenty pairs of cancerous tissues (CRC) and their adjacent noncancerous (control) tissues were subjected to quantitative reverse-transcription polymerase chain reaction analyses to examine mRNA levels of CUL1, CUL4A, and CUL4B. **p < 0.01, ***p < 0.001. (D–G) Kaplan-Meier survival curves. CRC tumor samples harboring higher and lower levels of CUL1 (D), CUL2 (E), CUL4A (F), and CUL4B (G) in The Cancer Genome Atlas database were analyzed to examine survival rates using Kaplan-Meier plots. (H) Immunohistochemistry (IHC) staining results. Three pairs of CRC and their adjacent noncancerous counterpart (control) tissues were subjected to IHC staining to examine protein levels of CUL1, CUL2, CUL4A, and CUL4B. One group of representative images is shown. Scale bars=50 mm. (I) Protein levels of cullins. The same sources of biopsies as in (H) were used for immunoblots to examine the protein levels of CUL1, CUL2, CUL4A, CUL4B, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
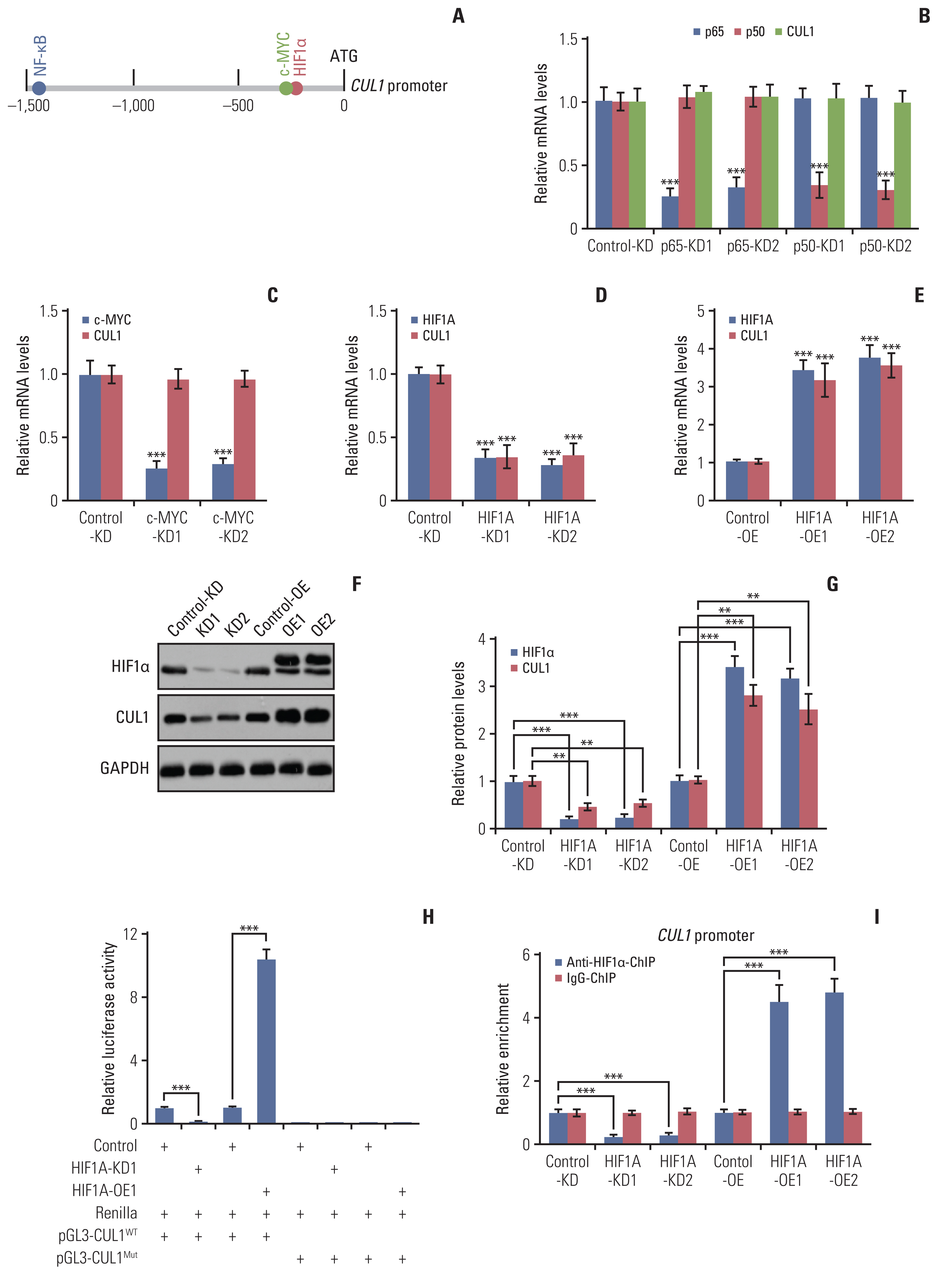
Fig. 2 Hypoxia-inducible factor 1α (HIF1α) specifically controlled the expression of CUL1. (A) The transcription factor binding sites on the promoter of CUL1. Three transcription factors (nuclear factor κB [NF-κB], c-MYC, and HIF1α) were predicted to bind to the promoter of CUL1 (1,500 bp length) and their binding sites are shown. (B) The effects of NF-κB subunits on the expression of CUL1. Total RNA isolated from Control-KD, p65-KD (#1 and #2), and p50-KD (#1 and #2) cells in the CCD-18Co background were used to detect the mRNA levels of p65, p50, and CUL1. (C) The effects of c-MYC on the expression of CUL1. Total RNA isolated from Control-KD and c-MYC-KD (#1 and #2) cells in the CCD-18Co background were used to detect the mRNA levels of c-MYC and CUL1. (D, E) The effects of HIF1α on the expression of CUL1. Total RNA isolated from Control-KD, HIF1a-KD (#1 and #2) (D), Control-OE, and HIF1A-OE (#1 and #2) cells (E) in the CCD-18Co background were used to detect the mRNA levels of HIF1A and CUL1. (F, G) The protein levels of CUL1 in HIF1A-KD and HIF1A-OE cells. Total cell extracts from cells (same as in D and E) were subjected to immunoblots to examine protein levels of HIF1α, CUL1, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (F). The protein signals were quantified and normalized to GAPDH (G). (H) The relative luciferase activities. The pGL4.3-pCUL1WT and pGL4.3-pCUL1Mut plasmids were co-transfected with Renilla into Control-KD, HIF1A-KD1, Control-OE, and HIF1A-OE1 cells in the CCD-18Co background. After 16 hours, the cells were harvested and used in dual-luciferase reporter assays. (I) The enrichment of HIF1α on the promoter of CUL1. Cells (as in D and E) were used for chromatin immunoprecipitation assays with anti-HIF1α and IgG (negative control). The input and output DNA were subjected to quantitative reverse-transcription polymerase chain reaction analyses to detect the enrichment of HIF1α and IgG on the promoter of CUL1. The relative enrichment of HIF1α and IgG in each sample was determined by normalizing to the levels in the Control-KD cells. **p < 0.01, ***p < 0.001.

Fig. 3 CUL1 assembled a complex with RING-box protein 1 (RBX1), S-phase kinase associated protein 1 (SKP1), and F-box/LRR-repeat protein 1 (FBXL1). (A) CUL1 pulled down RBX1, SKP1, and FBXL1 in vivo. Equal weights of three colorectal cancer (CRC) tumor tissues were mixed together and subjected to immunoprecipitation (IP) assays using IgG- or anti-CUL1–coupled protein A beads. The purified complexes and their corresponding inputs were probed with the antibodies shown in the figures. (B) The protein levels of RBX1, SKP1, and FBXL1 in CRC tumors. The same protein extracts as in Fig. 1I were subjected to immunoblots to examine protein levels of CUL1, RBX1, SKP1, FBXL1, and GAPDH. (C) SKP1 pulled down FBXL1 in vitro. CCD18-Co cells transfected with different combinations of plasmids, as indicated in the figures, were immunoprecipitated with Flag agarose. The input and output proteins were subjected to immunoblots and probed with anti-Flag and anti-Myc antibodies. (D) A schematic diagram of the SCFFBXL1 E3 ligase. (E) MTT assay results. The Control-KD, CUL1-KD (#1 and #2), and FBXL1-KD (#1 and #2) cells in the HT29 background were assayed for cell proliferation and cell viability at 1-day intervals with the MTT assay. * p < 0.05, **p < 0.01. (F) Cell migration results. The same cells as shown in (E) were used in the cell migration assays. The migrated cells were stained with 0.2% crystal violet. Scale bars=100 mm.
Fig. 4 The SCFFBXL1 E3 ligase ubiquitinated menin 1 (MEN1) in colorectal cancer (CRC) cells. (A) F-box/LRR-repeat protein 1 (FBXL1) pulled down MEN1 in vivo. The same protein lysates as in Fig. 3A were used for immunoprecipitation (IP) analyses using anti-FBXL1–or IgG-coupled protein A beads. The input and output proteins were probed with the antibodies shown in the figures. (B) FBXL1 pulled down MEN1 in vitro. Different combinations of plasmids (as shown in the figure) were co-transfected into CCD18-Co cells and then immunoprecipitated with anti-Flag agarose. The input and output proteins were probed with anti-Flag and anti-Myc antibodies. (C) The protein level of MEN1 in CRC tumors. The same protein samples as in Fig. 1I were subjected to western blotting to examine protein levels of MEN1 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH). (D) MTT assay results. The Control-OE and MEN1-OE (#1 and #2) cells in the HT29 background were assays for cell proliferation and cell viability at 1-day intervals with the MTT assay. * p < 0.05, **p < 0.01. (E) The SCFFBXL1 E3 ligase ubiquitinated MEN1 in vivo. The Control-KD, CUL1-KD, and FBXL1-KD cells were co-transfected with Flag-MEN1 and HA-ubiquitin (Ub) and then treated with 10 μM MG132 for 4 hours before harvest, followed by IP assays using anti-Flag agarose. The enriched proteins were probed with anti-HA antibody. The total MEN1 (FlagMEN1) was used as a loading control. (F) A schematic diagram of MEN1 ubiquitination by the SCFFBXL1 E3 ligase.
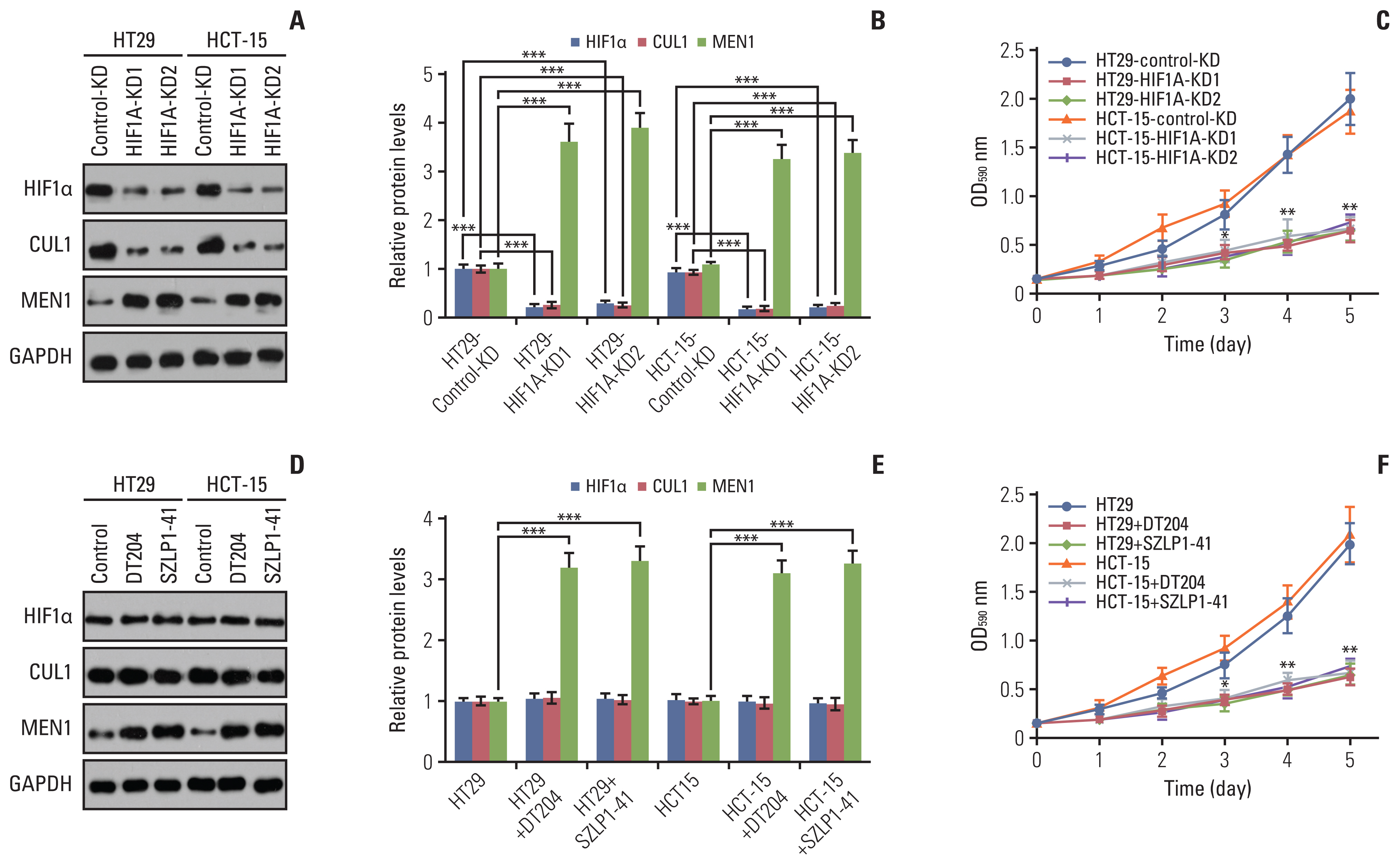
Fig. 5 The effects of knockdown of hypoxia-inducible factor 1α (HIF1α) and blockage of the SCFFBXL1 E3 ligase on menin 1 (MEN1) protein levels and colorectal cancer (CRC) cell growth. (A, B) Knockdown of HIF1α increased MEN1 protein level. The Control-KD and HIF1A-KD (#1 and #2) in both HT29 and HCT-15 backgrounds were used for immunoblots to examine the protein levels of HIF1α, CUL1, MEN1, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (A). The protein signals were quantified using Image J software and normalized to GAPDH (B). ***p < 0.001. (C) Knockdown of HIF1α inhibited CRC cell growth. Cells (as in A) were assayed for cell proliferation and cell viability determined at 1-day intervals with the MTT assay. (D, E) Blockage of SCFFBXL1 E3 ligase increased the MEN1 protein level. The HT29 and HCT cells were treated with 10 mm DT204 and 10 mm SZLP1-41 for 8 hours and then used for immunoblots to examine the protein levels of HIF1α, CUL1, MEN1, and GAPDH (D). The protein signals were quantified using Image J software and normalized to GAPDH (E). (F) Blockage of SCFFBXL1 E3 ligase inhibited CRC cell growth. Cells (as in D) were assayed for cell proliferation and cell viability determined at one-day intervals with the MTT assay. * p < 0.05, **p < 0.01, ***p < 0.001.
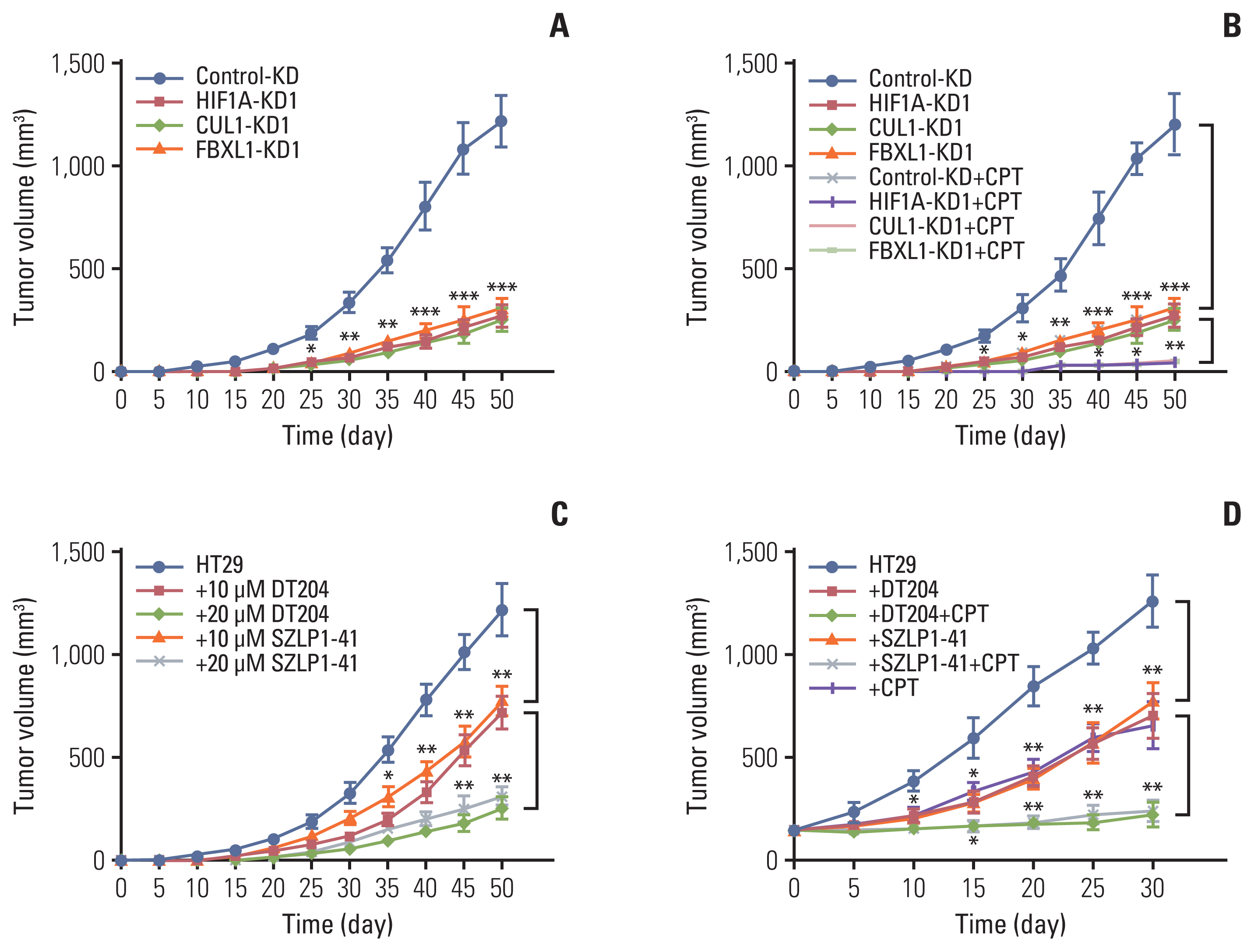
Fig. 6 Knockdown of hypoxia-inducible factor 1α (HIF1α), CUL1, or F-box/LRR-repeat protein 1 (FBXL1) or blockage of SCFFBXL1 E3 ligase significantly decreased tumor growth in vivo. (A) Knockdown of HIF1α, CUL1, or FBXL1 inhibited tumor growth. The Control-KD, HIF1AKD1, CUL1-KD1, and FBXL1-KD1 cells in the HT29 background were injected into nude mice. Tumor volumes were measured every 5 days. (B) Knockdown of HIF1a, CUL1, or FBXL1 increased the sensitivity of tumors to capecitabine (CPT). The same cells as in (A) were injected into nude mice, followed by injection of phosphate buffered saline (PBS) or CPT at 5-day intervals. Tumor volumes were measured every 5 days. (C) DT204 and SZLP1-411 inhibited tumor growth. The HT29 cells were injected into nude mice and amice were randomly divided into five groups, followed by injecting with PBS, DT204 (10 mm and 20 mm), and SZLP1-41 (10 mm and 20 mm) at 5-day intervals, respectively. Tumor volumes were also measured every 5 days. (D) DT204 and SZLP1-41 treatments increased the sensitivity of tumors to CPT. The HT29 cells were injected into nude mice and the mice injected with PBS, DT204, SZLP1-41, CPT, DT204+CPT, or SZLP1-41+CPT at 5-day intervals. Tumor volumes were measured every 5 days. *p < 0.05, **p < 0.01, ***p < 0.001.
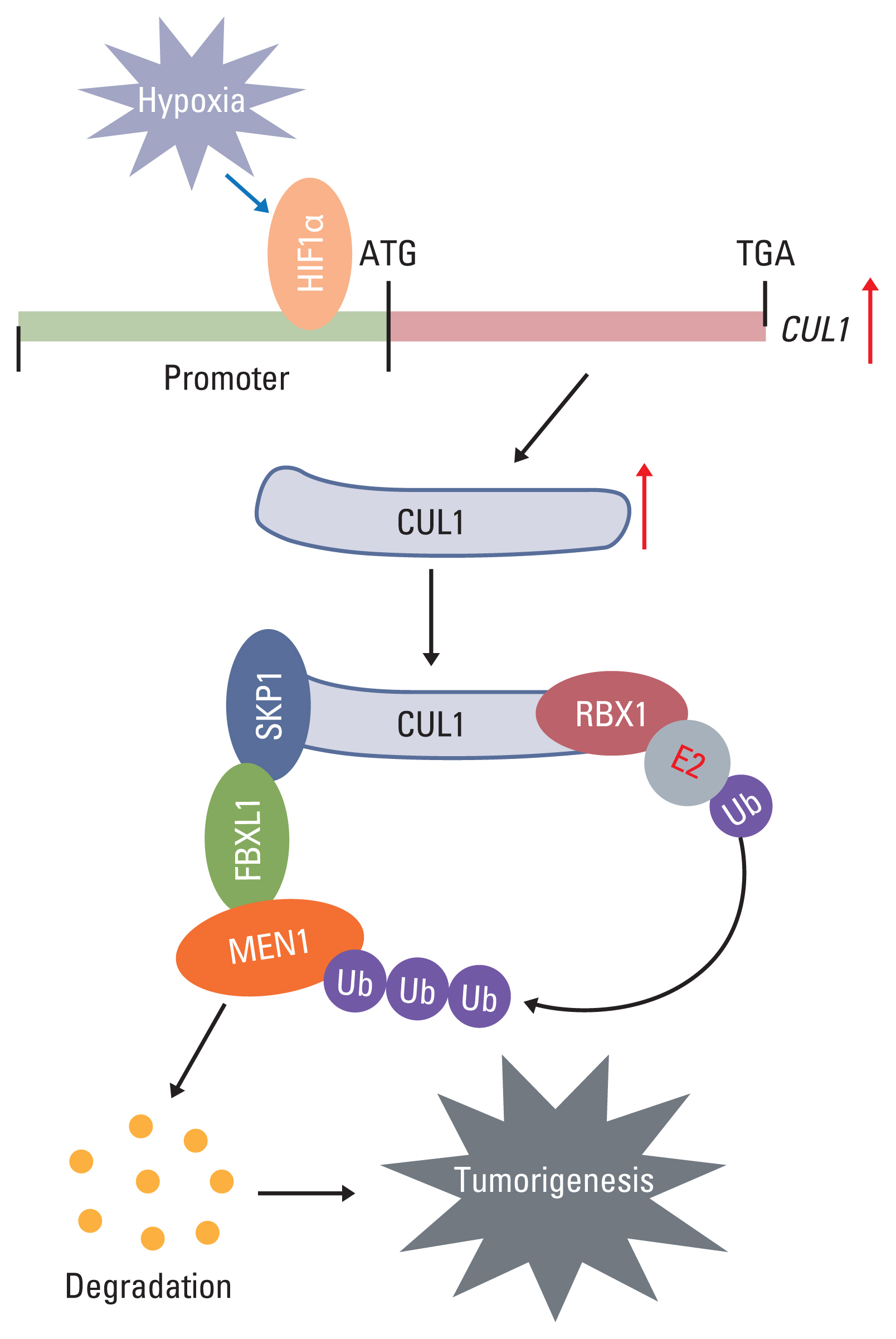
Fig. 7 A schematic diagram of SCFFBXL1E3 ligase-mediated ubiquitination of menin 1 (MEN1) in colorectal cancer tumorigenesis. Hypoxia enhances the binding of hypoxia-inducible factor 1α (HIF1α) onto the CUL1 promoter and activates expression of the CUL1 gene. The induced CUL1 recruits both RING-box protein 1 (RBX1) and S-phase kinase associated protein 1 (SKP1) to assemble a complex. RBX1 binds to the E2 ubiquitin (Ub)-conjugating enzyme, while SKP1 interacts with F-box/LRR-repeat protein 1 (FBXL1) to recognize MEN1 as a substrate. The degradation of MEN1 by the SCFFBXL1E3 ligase causes uncontrolled cell proliferation and results in tumorigenesis.
Reference
-
References
1. Rawla P, Sunkara T, Barsouk A. Epidemiology of colorectal cancer: incidence, mortality, survival, and risk factors. Prz Gastroenterol. 2019; 14:89–103.
Article2. Karimi S, Abdi A, Khatony A, Akbari M, Faraji A. Epidemiology of colorectal cancer and the risk factors in Kermanshah province-Iran 2009–2014. J Gastrointest Cancer. 2019; 50:740–3.
Article3. Yao N, Wang J, Cai Y, Yuan J, Wang H, Gong J, et al. Patterns of cancer screening, incidence and treatment disparities in China: protocol for a population-based study. BMJ Open. 2016; 6:e012028.
Article4. Huang Q, Figueiredo-Pereira ME. Ubiquitin/proteasome pathway impairment in neurodegeneration: therapeutic implications. Apoptosis. 2010; 15:1292–311.
Article5. Weathington NM, Mallampalli RK. Emerging therapies targeting the ubiquitin proteasome system in cancer. J Clin Invest. 2014; 124:6–12.
Article6. Brooks CL, Gu W. p53 regulation by ubiquitin. FEBS Lett. 2011; 585:2803–9.
Article7. Trotman LC, Wang X, Alimonti A, Chen Z, Teruya-Feldstein J, Yang H, et al. Ubiquitination regulates PTEN nuclear import and tumor suppression. Cell. 2007; 128:141–56.
Article8. Mulder MP, Witting K, Berlin I, Pruneda JN, Wu KP, Chang JG, et al. A cascading activity-based probe sequentially targets E1-E2-E3 ubiquitin enzymes. Nat Chem Biol. 2016; 12:523–30.
Article9. Chen Z, Sui J, Zhang F, Zhang C. Cullin family proteins and tumorigenesis: genetic association and molecular mechanisms. J Cancer. 2015; 6:233–42.
Article10. Zhao Y, Sun Y. Cullin-RING Ligases as attractive anti-cancer targets. Curr Pharm Des. 2013; 19:3215–25.
Article11. Soucy TA, Dick LR, Smith PG, Milhollen MA, Brownell JE. The NEDD8 conjugation pathway and its relevance in cancer biology and therapy. Genes Cancer. 2010; 1:708–16.
Article12. Liu H, Lu W, He H, Wu J, Zhang C, Gong H, et al. Inflammation-dependent overexpression of c-Myc enhances CRL4 (DCAF4) E3 ligase activity and promotes ubiquitination of ST7 in colitis-associated cancer. J Pathol. 2019; 248:464–75.13. Nelakurti DD, Pappula AL, Rajasekaran S, Miles WO, Petreaca RC. Comprehensive analysis of MEN1 mutations and their role in cancer. Cancers (Basel). 2020; 12:2616.
Article14. Matkar S, Thiel A, Hua X. Menin: a scaffold protein that controls gene expression and cell signaling. Trends Biochem Sci. 2013; 38:394–402.
Article15. Kamilaris CDC, Stratakis CA. Multiple endocrine neoplasia type 1 (MEN1): an update and the significance of early genetic and clinical diagnosis. Front Endocrinol (Lausanne). 2019; 10:339.
Article16. Li C, Xiao XQ, Qian YH, Zhou ZY. The CtBP1-p300-FOXO3a transcriptional complex represses the expression of the apoptotic regulators Bax and Bim in human osteosarcoma cells. J Cell Physiol. 2019; 234:22365–77.17. Majmundar AJ, Wong WJ, Simon MC. Hypoxia-inducible factors and the response to hypoxic stress. Mol Cell. 2010; 40:294–309.
Article18. Xie J, Jin Y, Wang G. The role of SCF ubiquitin-ligase complex at the beginning of life. Reprod Biol Endocrinol. 2019; 17:101.
Article19. Malek E, Abdel-Malek MA, Jagannathan S, Vad N, Karns R, Jegga AG, et al. Pharmacogenomics and chemical library screens reveal a novel SCF(SKP2) inhibitor that overcomes Bortezomib resistance in multiple myeloma. Leukemia. 2017; 31:645–53.
Article20. Chan CH, Morrow JK, Li CF, Gao Y, Jin G, Moten A, et al. Pharmacological inactivation of Skp2 SCF ubiquitin ligase restricts cancer stem cell traits and cancer progression. Cell. 2013; 154:556–68.
Article21. Penas C, Ramachandran V, Ayad NG. The APC/C ubiquitin ligase: from cell biology to tumorigenesis. Front Oncol. 2011; 1:60.
Article22. Galindo-Moreno M, Giraldez S, Limon-Mortes MC, Belmonte-Fernandez A, Reed SI, Saez C, et al. SCF(FBXW7)-mediated degradation of p53 promotes cell recovery after UV-induced DNA damage. FASEB J. 2019; 33:11420–30.
Article23. Lu Y, Li J, Cheng D, Parameswaran B, Zhang S, Jiang Z, et al. The F-box protein FBXO44 mediates BRCA1 ubiquitination and degradation. J Biol Chem. 2012; 287:41014–22.
Article24. Wu G, Xu G, Schulman BA, Jeffrey PD, Harper JW, Pavletich NP. Structure of a beta-TrCP1-Skp1-beta-catenin complex: destruction motif binding and lysine specificity of the SCF(beta-TrCP1) ubiquitin ligase. Mol Cell. 2003; 11:1445–56.25. Chen Z, Wang K, Hou C, Jiang K, Chen B, Chen J, et al. CRL4B(DCAF11) E3 ligase targets p21 for degradation to control cell cycle progression in human osteosarcoma cells. Sci Rep. 2017; 7:1175.
Article26. Chen L, Liu S, Tao Y. Regulating tumor suppressor genes: post-translational modifications. Signal Transduct Target Ther. 2020; 5:90.
Article27. Aubry A, Yu T, Bremner R. Preclinical studies reveal MLN4924 is a promising new retinoblastoma therapy. Cell Death Discov. 2020; 6:2.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Ubiquitin-Proteasome System and F-box Proteins in Pathogenic Fungi
- Phospholipase D2 promotes degradation of hypoxia-inducible factor-1alpha independent of lipase activity
- A Study on the Expression of p53 and nm23 Protein in the Colorectal Adenoma and Carcinoma
- Mannose Attenuates Colitis-Associated Colorectal Tumorigenesis by Targeting Tumor-Associated Macrophages
- The Role of MicroRNAs in Colorectal Cancer
