Deficiency of Follistatin-Like Protein 1 Accelerates the Growth of Breast Cancer Cells at Lung Metastatic Sites
- Affiliations
-
- 1Department of Human Anatomy, School of Basic Medical Sciences, Capital Medical University, Beijing, China. gy1003@ccmu.edu.cn
- 2Department of Breast Surgery, Cancer Institute and Hospital, Chinese Academy of Medical Sciences, Beijing, China.
- 3Beijing Key Laboratory of Cancer Invasion and Metastasis Research, Beijing, China.
- 4Cancer Institute, Capital Medical University, Beijing, China.
- 5Department of Breast Surgery, Cancer Hospital of Huanxing Chaoyang District Beijing, Beijing, China.
- 6Department of Clinical Laboratory, Cancer Hospital of Huanxing Chaoyang District Beijing, Beijing, China.
- KMID: 2421367
- DOI: http://doi.org/10.4048/jbc.2018.21.e43
Abstract
- PURPOSE
Follistatin-like protein 1 (FSTL1) is a secreted glycoprotein that has been shown to play a role in various types of cancer. However, the clinical significance and function of FSTL1 in breast cancer have not been reported. We investigated the role of FSTL1 in breast cancer in this study.
METHODS
Enzyme-linked immunosorbent assays, western blot analysis, and reverse transcription polymerase chain reaction were used to monitor the expression of FSTL1 in breast cancer tissue and in serum samples from breast cancer patients. We employed a 4T1 breast cancer model and Fstl1(+/−) mice for in vivo studies. Hematoxylin and eosin staining, western blot analysis, and RNA sequencing were used to analyze the effect of FSTL1 on primary tumor growth and lung metastasis.
RESULTS
We demonstrated that the expression of FSTL1 is reduced in both the breast cancer tissue and the serum of breast cancer patients. We showed that reduced levels of FSTL1 in serum correlate with elevated expression of Ki-67 and epidermal growth factor receptor (EGFR) in cancer tissues. Moreover, lowered expression of FSTL1 was associated with decreased survival in breast cancer patients. Experiments on the Fstl1(+/−) mouse model established that FSTL1 deficiency had no effect on primary tumor growth, but increased the lung metastases of breast cancer cells, resulting in reduced survival of tumor-bearing mice. RNA sequencing found significantly reduced expression of Egln3 and increased expression of EGFR in Fstl1(+/−) mice. Thus, our results suggest that FSTL1 may affect the expression of EGFR through Egln3, inhibiting the proliferation of breast cancer cells at lung metastatic sites.
CONCLUSION
In conclusion, we suggest a suppressor role of FSTL1 in breast cancer lung metastasis. Furthermore, FSTL1 may represent a potential prognostic biomarker and a candidate therapeutic target in breast cancer patients.
MeSH Terms
-
Animals
Blotting, Western
Breast Neoplasms*
Breast*
Enzyme-Linked Immunosorbent Assay
Eosine Yellowish-(YS)
Follistatin-Related Proteins*
Genes, Tumor Suppressor
Glycoproteins
Hematoxylin
Humans
Lung*
Mice
Neoplasm Metastasis
Polymerase Chain Reaction
Receptor, Epidermal Growth Factor
Reverse Transcription
Sequence Analysis, RNA
Eosine Yellowish-(YS)
Follistatin-Related Proteins
Glycoproteins
Hematoxylin
Receptor, Epidermal Growth Factor
Figure
-
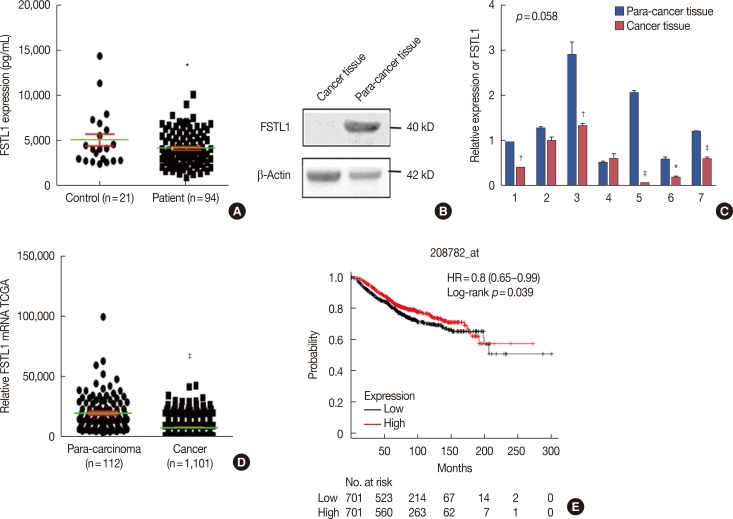
Figure 1 The expression of follistatin-like protein 1 (FSTL1) in breast cancer patients. (A) Serum levels of FSTL1 were determined by enzyme-linked immunosorbent assay in breast cancer patients (n=94) and healthy controls (n=21). (B) Western blot analysis of FSTL1 expression in cancer tissue and marched para-carcinoma tissue (n=4). (C) Real-time reverse transcription polymerase chain reaction analysis of eight paired breast carcinoma samples for the expression of FSTL1. (D) Relative expression of FSTL1 messenger RNA in cancer tissue (n=1,101) and marched para-carcinoma tissue (n=112) in breast cancer patients from the Cancer Genome Atlas (TCGA) cohort. (E) Kaplan-Meier survival analysis of public database (TCGA 2017) showed low expression of FSTL1 was associated with a worse overall survival of breast cancer patients. Each bar represents mean±SD for triplicate experiments.HR=hazard ratio. *p<0.05; †p<0.01; ‡p<0.001, unpaired t-test.
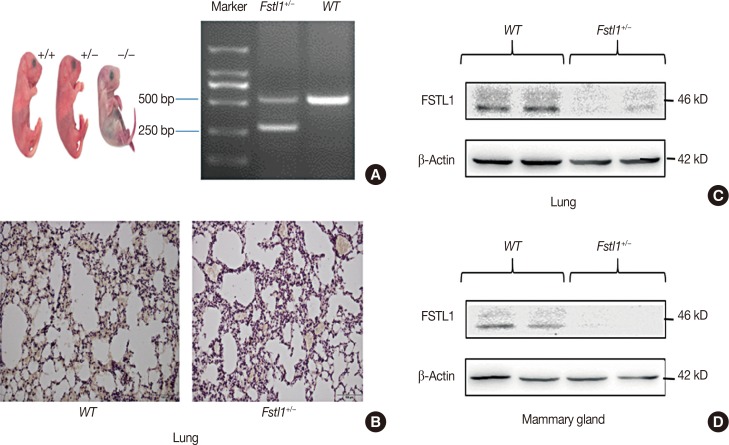
Figure 2 Generation of Fstl1+/− mice. (A) Genotyping of mice. (B) Histological analysis of follistatin-like protein 1 (FSTL1) in lung tissue of wild type and Fstl1+/− mice (n=3) by immunohistochemical (IHC) staining (IHC for FSTL1, ×400). (C) Western blot analysis of FSTL1 in lung tissue of WT and Fstl1+/− mice. (D) Western blot analysis of FSTL1 in mammary tissue of WT and Fstl1+/− mice.bp =base pair.
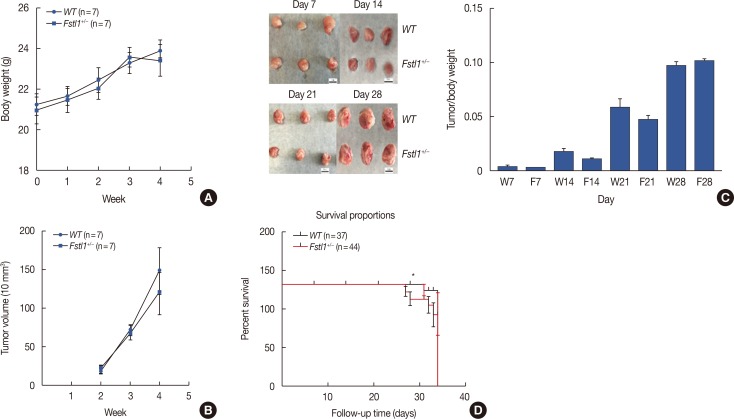
Figure 3 Growth of primary 4T1 tumors in wild type (WT) and Fstl1+/−mice. (A) Body weight was monitored in WT and Fstl1+/−mice for 4 weeks after the injection of 4T1 cancer cells (n=7). (B) Tumor volume was measured for 3 weeks (n=7). (C) The dissected tumors at day 7, 14, 21, and 28 (n=3). Tumors were dissected, and their weights and tumor volumes were measured. (D) Survival analysis of WT (n=37) and Fstl1+/−mice (n=44). Each bar represents the mean±SD for triplicate experiments.W=WT; Fstl1+/−. *p<0.05, unpaired t-test.
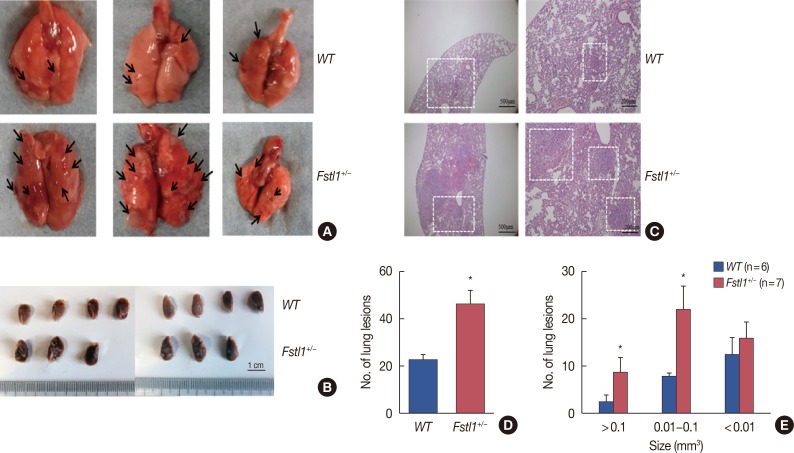
Figure 4 Increased lung metastasis in Fstl1+/−mice. (A) Thirty-one days after injection of 4T1 cancer cells, the mice were euthanized and analyzed for lung metastasis. The arrows were pointed to the lung metastasis in fresh lung tissue. (B) Thirty-one days after injection of 4T1 cancer cells, the mice were euthanized and analyzed for lung metastasis of formalin fixation. (C) Representative H&E stained images of the lung. (D) Number of lung metastatic foci were assessed in wild type (WT) (n=6) and Fstl1+/−mice (n=7). (E) Number and size of lung metastatic foci were assessed in WT (n=6) and Fstl1+/−mice (n=7). Each bar represents mean±SD for triplicate experiments.*p<0.05, unpaired t-test.
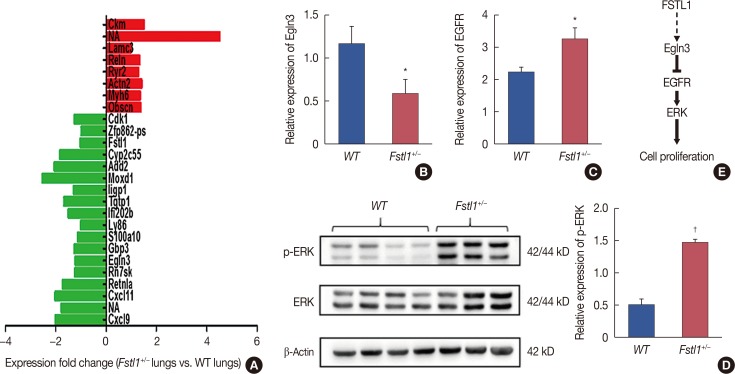
Figure 5 The epidermal growth factor receptor (EGFR) signaling in lung metastatic tumor tissue of Fstl1+/−mice. (A) Differential gene expression in lung metastatic tumor tissue of wild type (WT) (n=3) and Fstl1+/−mice (n=3). (B) Relative expression of Egln3 was decreased in lung metastatic tumor tissue of Fstl1+/−mice by real-time reverse transcription polymerase chain reaction (n=7). (C) Relative expression of EGFR was increased in lung metastatic tumor tissue of Fstl1+/−mice by transcription analysis (n=7). (D) Western blot analysis of phosphorylation of extracellular signal-regulated kinase (p-ERK) expression in lung metastatic tumor tissue of WT and Fstl1+/−mice. (E) Hypothetical model of follistatin-like protein 1 (FSTL1)-regulated cancer cell proliferation. Each bar represents the mean±SD for triplicate experiments.*p<0.05; †p<0.001, unpaired t-test.
Reference
-
1. Li J, Zhang BN, Fan JH, Pang Y, Zhang P, Wang SL, et al. A nation-wide multicenter 10-year (1999–2008) retrospective clinical epidemiological study of female breast cancer in China. BMC Cancer. 2011; 11:364. PMID: 21859480.
Article2. Peddi PF, Ellis MJ, Ma C. Molecular basis of triple negative breast cancer and implications for therapy. Int J Breast Cancer. 2012; 2012:217185. PMID: 22295242.
Article3. Rakha EA, Elsheikh SE, Aleskandarany MA, Habashi HO, Green AR, Powe DG, et al. Triple-negative breast cancer: distinguishing between basal and nonbasal subtypes. Clin Cancer Res. 2009; 15:2302–2310. PMID: 19318481.
Article4. Lehmann BD, Bauer JA, Chen X, Sanders ME, Chakravarthy AB, Shyr Y, et al. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J Clin Invest. 2011; 121:2750–2767. PMID: 21633166.
Article5. Shibanuma M, Mashimo J, Mita A, Kuroki T, Nose K. Cloning from a mouse osteoblastic cell line of a set of transforming-growth-factor-beta 1-regulated genes, one of which seems to encode a follistatin-related polypeptide. Eur J Biochem. 1993; 217:13–19. PMID: 7901004.6. Geng Y, Dong Y, Yu M, Zhang L, Yan X, Sun J, et al. Follistatin-like 1 (Fstl1) is a bone morphogenetic protein (BMP) 4 signaling antagonist in controlling mouse lung development. Proc Natl Acad Sci U S A. 2011; 108:7058–7063. PMID: 21482757.
Article7. Li D, Wang Y, Xu N, Wei Q, Wu M, Li X, et al. Follistatin-like protein 1 is elevated in systemic autoimmune diseases and correlated with disease activity in patients with rheumatoid arthritis. Arthritis Res Ther. 2011; 13:R17. PMID: 21303509.
Article8. Oshima Y, Ouchi N, Sato K, Izumiya Y, Pimentel DR, Walsh K. Follistatin-like 1 is an Akt-regulated cardioprotective factor that is secreted by the heart. Circulation. 2008; 117:3099–3108. PMID: 18519848.
Article9. Reddy SP, Britto R, Vinnakota K, Aparna H, Sreepathi HK, Thota B, et al. Novel glioblastoma markers with diagnostic and prognostic value identified through transcriptome analysis. Clin Cancer Res. 2008; 14:2978–2987. PMID: 18483363.
Article10. Su S, Parris AB, Grossman G, Mohler JL, Wang Z, Wilson EM. Up-regulation of follistatin-like 1 by the androgen receptor and melanoma antigen-a11 in prostate cancer. Prostate. 2017; 77:505–516. PMID: 27976415.
Article11. Lau MC, Ng KY, Wong TL, Tong M, Lee TK, Ming XY, et al. FSTL1 promotes metastasis and chemoresistance in esophageal squamous cell carcinoma through NFκB-BMP signaling cross-talk. Cancer Res. 2017; 77:5886–5899. PMID: 28883005.
Article12. Zhou X, Xiao X, Huang T, Du C, Wang S, Mo Y, et al. Epigenetic inactivation of follistatin-like 1 mediates tumor immune evasion in nasopharyngeal carcinoma. Oncotarget. 2016; 7:16433–16444. PMID: 26918942.
Article13. Zhao W, Han HB, Zhang ZQ. Suppression of lung cancer cell invasion and metastasis by connexin43 involves the secretion of follistatin-like 1 mediated via histone acetylation. Int J Biochem Cell Biol. 2011; 43:1459–1468. PMID: 21718795.
Article14. Chan QK, Ngan HY, Ip PP, Liu VW, Xue WC, Cheung AN. Tumor suppressor effect of follistatin-like 1 in ovarian and endometrial carcinogenesis: a differential expression and functional analysis. Carcinogenesis. 2009; 30:114–121. PMID: 18796737.15. Hodgson G, Hager JH, Volik S, Hariono S, Wernick M, Moore D, et al. Genome scanning with array CGH delineates regional alterations in mouse islet carcinomas. Nat Genet. 2001; 29:459–464. PMID: 11694878.
Article16. Liu Y, Han X, Yu Y, Ding Y, Ni C, Liu W, et al. A genetic polymorphism affects the risk and prognosis of renal cell carcinoma: association with follistatin-like protein 1 expression. Sci Rep. 2016; 6:26689. PMID: 27225192.
Article17. Hatzimichael E, Dasoula A, Shah R, Syed N, Papoudou-Bai A, Coley HM, et al. The prolyl-hydroxylase EGLN3 and not EGLN1 is inactivated by methylation in plasma cell neoplasia. Eur J Haematol. 2010; 84:47–51. PMID: 19737309.
Article18. Garvalov BK, Foss F, Henze AT, Bethani I, Gräf-Höchst S, Singh D, et al. PHD3 regulates EGFR internalization and signalling in tumours. Nat Commun. 2014; 5:5577. PMID: 25420589.
Article19. Henze AT, Garvalov BK, Seidel S, Cuesta AM, Ritter M, Filatova A, et al. Loss of PHD3 allows tumours to overcome hypoxic growth inhibition and sustain proliferation through EGFR. Nat Commun. 2014; 5:5582. PMID: 25420773.
Article20. Meng X, Zheng R, Zhang Y, Qiao M, Liu L, Jing P, et al. An activated sympathetic nervous system affects white adipocyte differentiation and lipolysis in a rat model of Parkinson's disease. J Neurosci Res. 2015; 93:350–360. PMID: 25257318.
Article21. Su Y, Loos M, Giese N, Hines OJ, Diebold I, Görlach A, et al. PHD3 regulates differentiation, tumour growth and angiogenesis in pancreatic cancer. Br J Cancer. 2010; 103:1571–1579. PMID: 20978507.
Article22. Xue J, Li X, Jiao S, Wei Y, Wu G, Fang J. Prolyl hydroxylase-3 is down-regulated in colorectal cancer cells and inhibits IKKbeta independent of hydroxylase activity. Gastroenterology. 2010; 138:606–615. PMID: 19786027.23. Bae K, Park KE, Han J, Kim J, Kim K, Yoon KA. Mitotic cell death caused by follistatin-like 1 inhibition is associated with up-regulated Bim by inactivated Erk1/2 in human lung cancer cells. Oncotarget. 2016; 7:18076–18084. PMID: 26716515.
Article24. Shi DL, Shi GR, Xie J, Du XZ, Yang H. MicroRNA-27a inhibits cell migration and invasion of fibroblast-like synoviocytes by targeting follistatin-like protein 1 in rheumatoid arthritis. Mol Cells. 2016; 39:611–618. PMID: 27498552.
Article25. Tsou PS, Wren JD, Amin MA, Schiopu E, Fox DA, Khanna D, et al. Histone deacetylase 5 is overexpressed in scleroderma endothelial cells and impairs angiogenesis via repression of proangiogenic factors. Arthritis Rheumatol. 2016; 68:2975–2985. PMID: 27482699.
Article26. Witton CJ, Reeves JR, Going JJ, Cooke TG, Bartlett JM. Expression of the HER1-4 family of receptor tyrosine kinases in breast cancer. J Pathol. 2003; 200:290–297. PMID: 12845624.
Article27. An J, Wang L, Zhao Y, Hao Q, Zhang Y, Zhang J, et al. Effects of FSTL1 on cell proliferation in breast cancer cell line MDA MB 231 and its brain metastatic variant MDA MB 231 BR. Oncol Rep. 2017; 38:3001–3010. PMID: 29048681.28. Gao H, Chakraborty G, Zhang Z, Akalay I, Gadiya M, Gao Y, et al. Multi-organ site metastatic reactivation mediated by non-canonical discoidin domain receptor 1 signaling. Cell. 2016; 166:47–62. PMID: 27368100.
Article29. Miyabe M, Ohashi K, Shibata R, Uemura Y, Ogura Y, Yuasa D, et al. Muscle-derived follistatin-like 1 functions to reduce neointimal formation after vascular injury. Cardiovasc Res. 2014; 103:111–120. PMID: 24743592.
Article30. Trojan L, Schaaf A, Steidler A, Haak M, Thalmann G, Knoll T, et al. Identification of metastasis-associated genes in prostate cancer by genetic profiling of human prostate cancer cell lines. Anticancer Res. 2005; 25(1A):183–191. PMID: 15816537.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Influence of Activin and Follistatin on Expression of Insulin-like Growth Factor (IGF)-I, II, Insulin-like Growth Factor Binding Protein (IGFBP)-1, 2, and 3 mRNA in Cultured Mouse Granulosa Cells
- Advances in Clinically Relevant Metastatic Breast Cancer Models
- Clinicopathological Study of 191 Cases of Metastatic Skin Cancers from Solid Cancer Diagnosed at the Department of Dermatology in a Tertiary Referral Cancer Center
- Multiple Cutaneous Edema and Infiltration of Signet-ring Cells in the Lymphatics as an Initial Manifestation of Metastatic Gastric Adenocarcinoma
- Analysis of PIK3CA Mutation Concordance and Frequency in Primary and Different Distant Metastatic Sites in Breast Cancer
