Heterogeneous Nuclear Ribonucleoprotein A2B1 Exerts a Regulatory Role in Lipopolysaccharide-stimulated 38B9 B Cell Activation
- Affiliations
-
- 1Department of Bioactive Material Sciences and Institute of Bioactive Materials, Chonbuk National University, Jeonju 54896, Korea. yongsuk@jbnu.ac.kr omfs1109@korea.ac.kr
- 2NBM Inc., Wanju 55322, Korea.
- 3Department of Molecular Biology and Institute for Molecular Biology and Genetics, Chonbuk National University, Jeonju 54896, Korea.
- 4Department of Dentistry, Graduate School of Medicine, Korea University, Seoul 02841, Korea. yongsuk@jbnu.ac.kr omfs1109@korea.ac.kr
- 5Department of Oral Maxillofacial Surgery, Korea University Ansan Hospital, Ansan 15355, Korea.
- KMID: 2400629
- DOI: http://doi.org/10.4110/in.2017.17.6.437
Abstract
- Major histocompatibility complex (MHC) class II molecules, which are recognized for their primary function of presenting an antigen to the T cell receptor, are involved in various signaling pathways in B cell activation. We identified heterogeneous nuclear ribonucleoprotein (hnRNP) A2B1 as an MHC class II molecule-associated protein involved in MHC class II-mediated signal transduction in lipopolysaccharide (LPS)-stimulated 38B9 B cells. Although the function of hnRNP A2B1 in the nucleus is primarily known, the level of hnRNP A2B1 in the cytoplasm was increased in LPS-stimulated 38B9 cells, while it was not detected in the cytoplasm of non-treated 38B9 cells. The silencing of hnRNP A2B1 expression using siRNA disturbed B cell maturation by regulation of mitogen-activated protein kinase signaling, NF-κB activation, and protein kinase B activation. These results suggest that hnRNP A2B1 is associated with MHC class II molecules and is involved in B cell activation signaling pathways in LPS-stimulated 38B9 cells.
Keyword
MeSH Terms
-
B-Lymphocytes
Cytoplasm
Heterogeneous-Nuclear Ribonucleoproteins*
Major Histocompatibility Complex
Protein Kinases
Proto-Oncogene Proteins c-akt
Receptors, Antigen, T-Cell
RNA, Small Interfering
Signal Transduction
Heterogeneous-Nuclear Ribonucleoproteins
Protein Kinases
Proto-Oncogene Proteins c-akt
RNA, Small Interfering
Receptors, Antigen, T-Cell
Figure
-
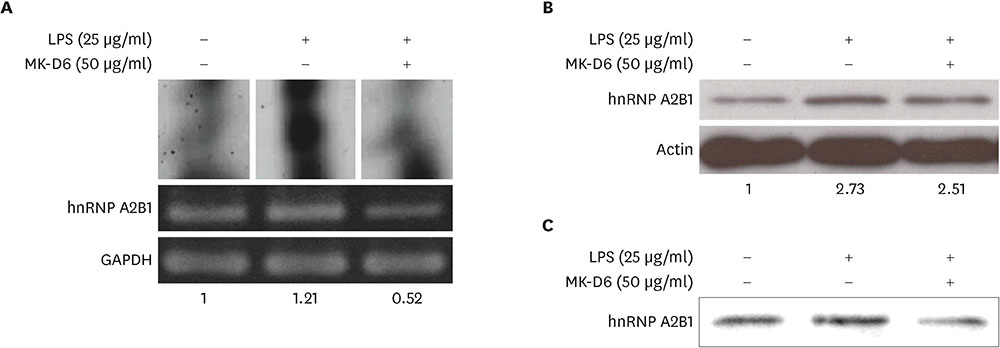
Figure 1 Identification of hnRNP A2B1 as an MHC class II-associated protein in 38B9 cells. (A) Partial 2-DE image of hnRNP A2B1 (upper panel) and the RT-PCR product of hnRNP A2B1 mRNA (lower panel) prepared from 38B9 cells in the presence or absence of LPS and anti-MHC class II Ab. MHC class II-associated proteins were prepared by immunoprecipitation from whole-cell lysates using an anti-MHC class II Ab and separated by 2-DE as described in the Materials and Methods section. GAPDH was used as an internal loading control for RT-PCR. Molecular mass and isoelectric point of the candidate protein included in the spot were calculated as 37.4 kDa and 8.97, respectively, using AxPASy inline tool and candidate protein was expected to be hnRNP A2B1 based on mass analysis data. (B) Western blot analysis of hnRNP A2B1 protein present in 38B9 cells after the indicated treatments. Actin was used as an internal loading control. (C) Whole-cell lysates from 38B9 cells were immunoprecipitated using the anti-MHC class II Ab after the indicated treatments and immunoblotted using the hnRNP A2B1 Ab. The numbers under each gel picture represent the RQ. Representative data from at least 3 independent experiments are presented.
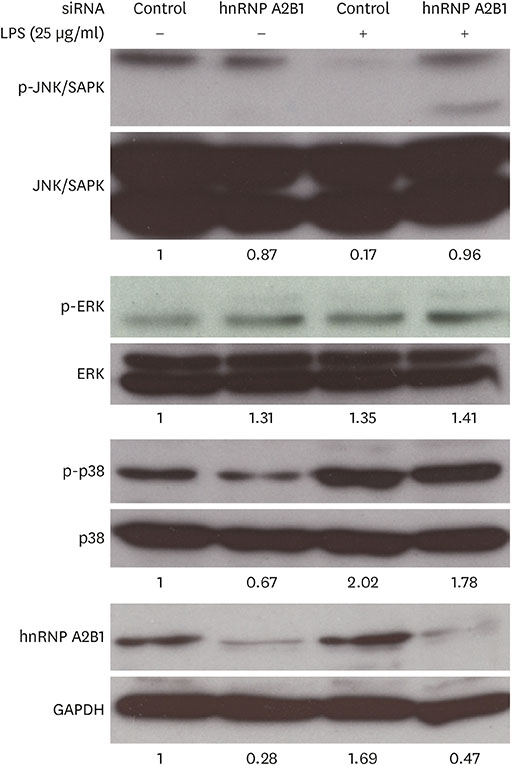
Figure 2 Effect of hnRNP A2B1 on the activation of components of the MAP kinase signaling pathway in 38B9 cells. The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS for 30 min at 37°C. Western blot analyses of the levels of total and p-ERK, p38, and JNK/SAPK in 38B9 cells following the indicated treatments after knocking down hnRNP A2B1 as described in the Materials and Methods section. Actin was used as an internal loading control. The numbers under each gel picture represent the RQ. Representative data from at least 3 independent experiments are presented. SAPK, stress-activated protein kinase.
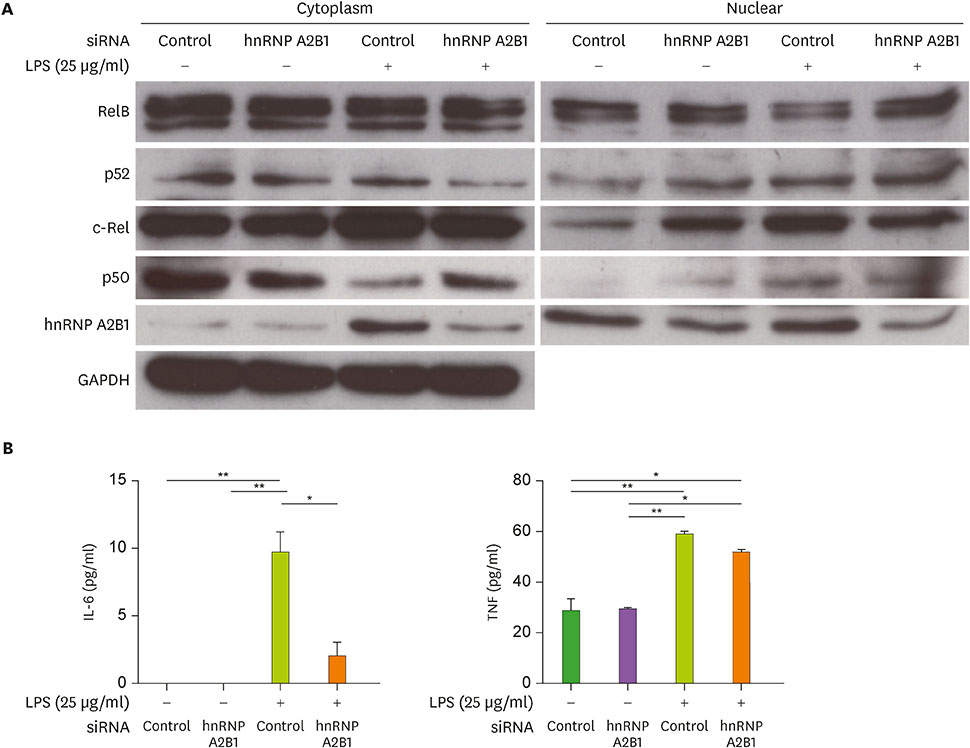
Figure 3 Effect of hnRNP A2B1 siRNA treatment on LPS-mediated NF-κB activation and the expression of pro-inflammatory cytokines in 38B9 cells. (A) The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS for 30 min at 37°C, and proteins in the cytoplasmic and nuclear fractions were analyzed by western blotting. GAPDH was used as an internal control. Representative data from 2 independent experiments are presented. (B) The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS for 12 h at 37°C, and cell culture supernatants were harvested. The levels of IL-6 (left panel) and TNF (right panel) in the culture supernatants were analyzed as described in the Materials and Methods section. Data are presented as means±standard deviation of duplicate determinations, and a representative result from 2 independent experiments is shown. TNF, tumor necrosis factor. *p<0.05 and **p<0.01 indicate significant differences between groups.

Figure 4 Effect of hnRNP A2B1 siRNA treatment on components of the Akt signaling pathway in 38B9 cells. The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS at 37°C for 30 min. Western blot analyses of the expression and activation of Akt, GSK3β, and Mcl-1 were performed in cells after the indicated treatments. Actin was used as an internal loading control for western blot analyses. The numbers under each gel picture represent the RQ. Representative data from at least 3 independent experiments are presented.
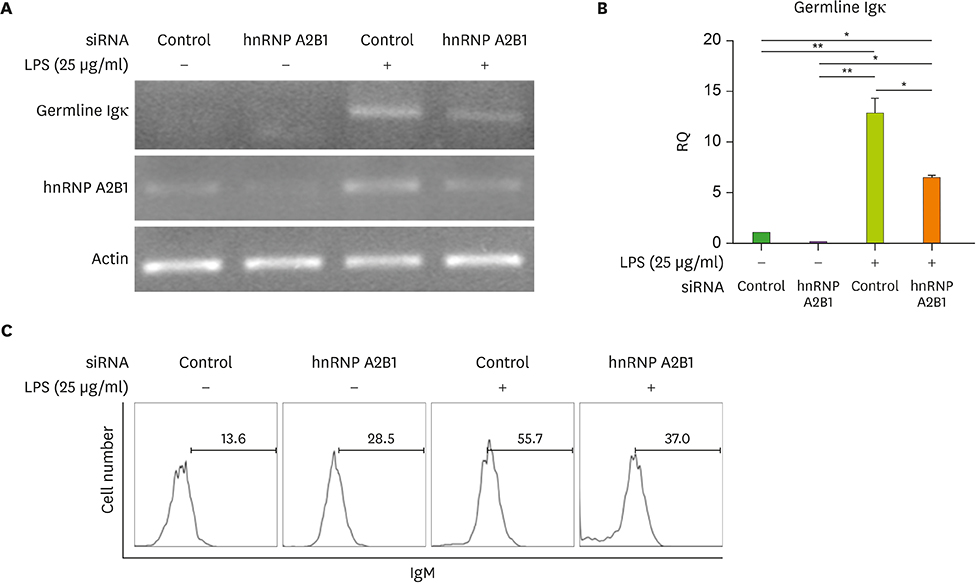
Figure 5 Effect of hnRNP A2B1 silencing on the differentiation of LPS-treated 38B9 cells. The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS at 37°C for 12 h. Expression of the germline Igκ gene in 38B9 cells after the indicated treatments was determined by (A) RT-PCR and (B) quantitative real-time PCR. In addition, the level of surface IgM expression was determined by flow cytometry. Representative data from at least 3 independent experiments are shown. *p<0.05 and **p<0.01 indicate significant differences between groups.
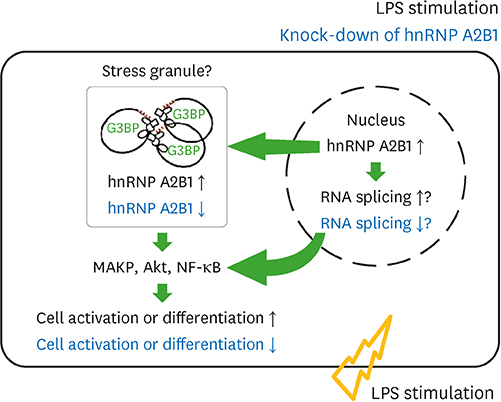
Figure 6 Schematic diagram outlining possible regulatory mechanism of cellular responses by hnRNP A2B1 in LPS-stimulated 38B9 cells. LPS stimulation of 38B9 cells increases the level of hnRNP A2B1 protein and may accumulate in stress granule in cytoplasm, which, in turn, activates cellular signaling pathways to activate 38B9 cells. These signaling pathways are suppressed by knocking-down of hnRNP A2B1.
Reference
-
1. Zhang W, Liu HT. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002; 12:9–18.
Article2. Huang G, Shi LZ, Chi H. Regulation of JNK and p38 MAPK in the immune system: signal integration, propagation and termination. Cytokine. 2009; 48:161–169.
Article3. Khiem D, Cyster JG, Schwarz JJ, Black BL. p38 MAPK-MEF2C pathway regulates B-cell proliferation. Proc Natl Acad Sci USA. 2008; 105:17067–17072.4. Limon JJ, Fruman DA. Akt and mTOR in B cell activation and differentiation. Front Immunol. 2012; 3:228.
Article5. Huang H, Tindall DJ. Dynamic FoxO transcription factors. J Cell Sci. 2007; 120:2479–2487.
Article6. Robinson JH, Delvig AA. Diversity in MHC class II antigen presentation. Immunology. 2002; 105:252–262.
Article7. Al-Daccak R, Mooney N, Charron D. MHC class II signaling in antigen-presenting cells. Curr Opin Immunol. 2004; 16:108–113.
Article8. Liu X, Zhan Z, Li D, Xu L, Ma F, Zhang P, Yao H, Cao X. Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining activation of the kinase Btk. Nat Immunol. 2011; 12:416–424.
Article9. Schneppenheim J, Loock AC, Hüttl S, Schweizer M, Lüllmann-Rauch R, Oberg HH, Arnold P, Lehmann CH, Dudziak D, Kabelitz D, et al. The influence of MHC class II on B cell defects induced by invariant chain/CD74 N-terminal fragments. J Immunol. 2017; 199:172–185.
Article10. Schneppenheim J, Hüttl S, Kruchen A, Fluhrer R, Müller I, Saftig P, Schneppenheim R, Martin CL, Schröder B. Signal-peptide-peptidase-like 2a is required for CD74 intramembrane proteolysis in human B cells. Biochem Biophys Res Commun. 2014; 451:48–53.
Article11. Yang HY, Kim J, Lee KY, Jang YS. Rac/ROS-related protein kinase C and phosphatidylinositol-3-kinase signaling are involved in a negative regulating cascade in B cell activation by antibody-mediated cross-linking of MHC class II molecules. Mol Immunol. 2010; 47:706–712.
Article12. Yang HY, Kim J, Kim SH, Choe CH, Jang YS. Pro-IL-16 is associated with MHC class II-mediated negative regulation of mouse resting B cell activation through MAP kinases, NF-κB and Skp2-dependent p27kip regulation. Scand J Immunol. 2013; 77:177–186.
Article13. Schlissel MS, Baltimore D. Activation of immunoglobulin kappa gene rearrangement correlates with induction of germline kappa gene transcription. Cell. 1989; 58:1001–1007.
Article14. Kim J, Kim HR, Lee JC, Jang YS. Involvement of ERK, p38 MAP kinase, and PKC in MHC class II-mediated signal transduction in a resting B cell line. Biochem Biophys Res Commun. 2002; 291:139–145.
Article15. Choi HS, Lee HM, Jang YJ, Kim CH, Ryu CJ. Heterogeneous nuclear ribonucleoprotein A2/B1 regulates the self-renewal and pluripotency of human embryonic stem cells via the control of the G1/S transition. Stem Cells. 2013; 31:2647–2658.
Article16. Gerondakis S, Siebenlist U. Roles of the NF-κB pathway in lymphocyte development and function. Cold Spring Harb Perspect Biol. 2010; 2:a000182.
Article17. Alt FW, Yancopoulos GD, Blackwell TK, Wood C, Thomas E, Boss M, Coffman R, Rosenberg N, Tonegawa S, Baltimore D. Ordered rearrangement of immunoglobulin heavy chain variable region segments. EMBO J. 1984; 3:1209–1219.
Article18. Krecic AM, Swanson MS. hnRNP complexes: composition, structure, and function. Curr Opin Cell Biol. 1999; 11:363–371.
Article19. Piñol-Roma S, Dreyfuss G. hnRNP proteins: localization and transport between the nucleus and the cytoplasm. Trends Cell Biol. 1993; 3:151–155.
Article20. Hernández-Díaz I, Giraldez T, Morales S, Hernandez G, Salido E, Canessa CM, Alvarez de la Rosa D. Heterogeneous nuclear ribonucleoprotein A2/B1 is a tissue-specific aldosterone target gene with prominent induction in the rat distal colon. Am J Physiol Gastrointest Liver Physiol. 2013; 304:G122–G131.
Article21. Hayashi EA, Akira S, Nobrega A. Role of TLR in B cell development: signaling through TLR4 promotes B cell maturation and is inhibited by TLR2. J Immunol. 2005; 174:6639–6647.
Article22. Feng B, Cheng S, Pear WS, Liou HC. NF-κB inhibitor blocks B cell development at two checkpoints. Med Immunol. 2004; 3:1.23. Faes S, Dormond O. PI3K and AKT: unfaithful partners in cancer. Int J Mol Sci. 2015; 16:21138–21152.
Article24. Vikström IB, Slomp A, Carrington EM, Moesbergen LM, Chang C, Kelly GL, Glaser SP, Jansen JH, Leusen JH, Strasser A, et al. MCL-1 is required throughout B-cell development and its loss sensitizes specific B-cell subsets to inhibition of BCL-2 or BCL-XL. Cell Death Dis. 2016; 7:e2345.
Article25. Shilo A, Ben Hur V, Denichenko P, Stein I, Pikarsky E, Rauch J, Kolch W, Zender L, Karni R. Splicing factor hnRNP A2 activates the Ras-MAPK-ERK pathway by controlling A-Raf splicing in hepatocellular carcinoma development. RNA. 2014; 20:505–515.
Article26. Kim HJ, Kim NC, Wang YD, Scarborough EA, Moore J, Diaz Z, MacLea KS, Freibaum B, Li S, Molliex A, et al. Mutations in prion-like domains in hnRNPA2B1 and hnRNPA1 cause multisystem proteinopathy and ALS. Nature. 2013; 495:467–473.
Article27. Lemieux B, Blanchette M, Monette A, Mouland AJ, Wellinger RJ, Chabot B. A function for the hnRNP A1/A2 proteins in transcription elongation. PLoS One. 2015; 10:e0126654.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Antineuroinflammatory Effects of 7,3’,4’-Trihydroxyisoflavone in Lipopolysaccharide-Stimulated BV2 Microglial Cells through MAPK and NF-κB Signaling Suppression
- The Regulatory Mechanism of Src Familly Kinase in Lipopolysaccharide (LPS) induced HF-kappaB Activation Pathway
- Phosphatidylinositol-3-kinase Inhibitor Enhances Nitric Oxide Synthesis and Apoptosis in LPS-Stimulated Macrophages
- Shikonin Isolated from Lithospermum erythrorhizon Downregulates Proinflammatory Mediators in Lipopolysaccharide-Stimulated BV2 Microglial Cells by Suppressing Crosstalk between Reactive Oxygen Species and NF-kappaB
- Effect of Modulation of hnRNP L Levels on the Decay of bcl-2 mRNA in MCF-7 Cells
